
Представьте, что сегодня утром вас разбудил непонятный звук, исходящий от странного маленького прямоугольника с яркими надписями «отложить» и «выключить». Рука сама собой потянулась и нажала на слово «выключить» и странный звук прекратился. Вы окончательно открыли глаза и обнаружили себя в незнакомом помещении. Вы встали, опустили ноги вниз и с ужасом нащупали ими непонятное волосатое существо, глядящее на вас двумя огромными глазами. В смятении вы вежливо его обошли и направились к дырке в стене напротив. За ней расстилалось бессчетное множество огромных коробок, между которыми быстро перемещались какие-то тарахтящие коробки поменьше. Вы захотели рассмотреть их поближе, но сильно ударились лбом о невидимую преграду. От боли и страха вы опустились на пол и обхватили голову руками, не понимая, что вокруг происходит и кто, собственно, вы такие. Так, скорее всего, начинался бы каждый ваш день, если бы по несчастливому стечению обстоятельств вы полностью утратили механизмы функционирования долговременной памяти. Удручающая картина, не правда ли?
Действительно, способность фиксировать, сохранять и использовать информацию, полученную в ходе пережитого опыта, представляет собой одно из важнейших эволюционных приобретений нашего вида. Совокупность наших воспоминаний, по сути, и определяет то, кто мы есть. Мы знаем, как нас зовут, какого мы пола, сколько нам лет, где мы живем, как выглядим мы и наши родители. То, как мы реагируем на различные жизненные ситуации, зависит от того, как мы действовали в аналогичных ситуациях в прошлом. Все это утратится, потеряй мы долговременную память.
Как некоторые, наверное, уже подметили, в приведенном в начале статьи примере, произошла утрата не всей долговременной памяти, а только одной ее составляющей – эксплицитной или, как ее еще называют, декларативной. Память не представляет из себя единый процесс, происходящий в каких-то одних структурах мозга. Память – это скорее совокупность процессов, каждый из которых приспособлен для сохранения какого-то определенного вида информации на какой-то определенный промежуток времени. Иерархическая схема различных типов памяти представлена на рис. 1.
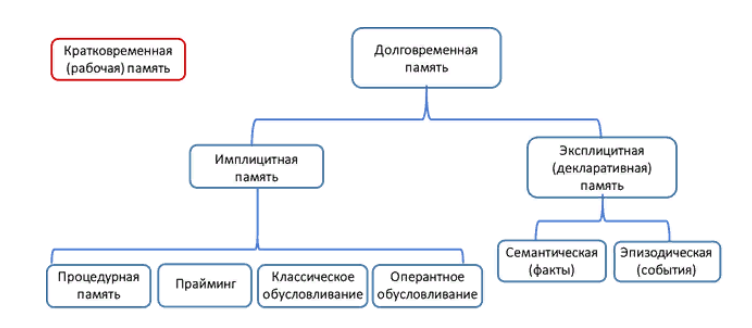
Рис. 1. Схема типов памяти
Очевидно, что не всю информацию необходимо запомнить на всю жизнь. Действительно, нам незачем запоминать два числа, которые нам нужно было сложить на контрольной по математике в 3-м классе. Подобного рода информация, необходимая для решения сиюминутных задач, «задерживается» в сознании совсем недолго и обрабатывается системой кратковременной (или рабочей) памяти. Данный домен памяти по своим нейробиологическим механизмам не имеет ничего общего с механизмами долговременной памяти и представляет собой отдельную сложную систему. В связи с этим вопросы, касающиеся функционирования кратковременной памяти, разбираться в данном материале не будут.
Вернемся к нашей контрольной по математике. Да, мы не помним, какие именно числа мы складывали, но мы помним, как вообще складываются числа и, возможно, помним саму эту контрольную, как выглядел школьный класс и, если вам очень повезло с памятью, недовольное лицо соседа по парте. За сохранение информации, которую мы можем вспомнить спустя много месяцев или лет, отвечает система долговременной памяти. Информацию, требующую длительного хранения, условно можно разделить на две большие группы. Информация первого рода отвечает на вопрос «что?» Что мы делали во время прошлого отпуска? Что нам вчера сказала подруга? Что лежит у нас в шкафу? Если мы попробуем ответить на эти вопросы, у нас в голове сразу же возникнет некое воспоминание, обладающее чувственными характеристиками. То есть, отвечая на последний вопрос, мы вспомним, как выглядит наш шкаф, как скрипят и гремят его дверцы, какова на ощупь его поверхность и как пахнет дерево, из которого он сделан. Подобная информация, составленная из совокупности наших чувственных ощущений, носит название эпизодической, то есть, содержит информацию о каком-то конкретном, пусть даже и повторяющемся, эпизоде из нашей жизни.
А теперь представьте, что нас спросили, что такое шкаф. Скорее всего мы ответим каким-то определением. Мы скажем, что шкаф – это предмет мебели для хранения вещей, или что-то наподобие этого. Да, отвечая на этот вопрос мы, скорее всего, тоже вспомним свой шкаф или представим себе некий усредненный абстрактный шкаф. Но в нашей голове появится также и определение этого слова, усредненное описание всех шкафов вообще, не связанное с восприятием через органы чувств. Информация такого рода носит название семантической и содержит понятия и их смысловые характеристики. И семантическая, и эпизодическая информация отвечает на вопрос «что?» и объединяется в единую систему эксплицитной, или декларативной памяти.
Однако для нормальной жизни нам необходимо не только помнить, что когда-либо происходило и что какое-нибудь понятие означает. Мы должны помнить, и как нам взаимодействовать с объектами, которые мы знаем. Как кататься на велосипеде? Как разговаривать с начальником? Как открыть этот надоевший шкаф? Да, прочитав эти вопросы сейчас и сконцентрировав свое внимание на них, у нас, скорее всего, возникнут в сознании некие образные воспоминания о конкретных событиях. Но если перед нами встанут эти проблемы в обычной рутинной жизни, мы вряд ли будем вспоминать прошлое, а просто сделаем все, как надо. Если нам понадобится открыть шкаф, мы не будем проговаривать последовательность действий про себя и не будем вспоминать, как открывали его в самый первый раз после покупки. Мы просто возьмемся за ручку и потянем в нужную сторону. Эту память, сохраняющую категорию информации, неосознаваемой нами в течение обычных процессов мышления, но критически важной для адаптации нас к внешней среде и корректирующей наши действия, называют имплицитной.
Существует несколько разных видов имплицитной памяти. В первую очередь, это процедурная память, позволяющая нам не учиться каждый раз каким-то сложным действиям, а делать их, что называется, «на автомате». К системе имплицитной памяти относится также известный эффект прайминга, заключающийся в усилении реагирования на определенные стимулы, если до этого мы уже были под действием этого же или даже лишь ассоциативно связанного стимула. К примеру, мы быстрее заметим среди бессвязного набора слов слово «пчела», если до этого мы прочитали слово «мёд». Исследователи включают в систему имплицитной памяти даже классическое павловское и оперантное обусловливание. Под первым понимают появление реакции не на сам первоначальный стимул, а на ассоциативно связанный с ним. Все мы помним про Павлова и его голодных собак, но и нас дверь с табличкой «стоматологический кабинет» может порой заставить поежиться, даже если нам туда и не надо. Под оперантным же обусловливанием понимают изменение вероятности определенной реакции на стимул в зависимости от типа подкрепления. Положительное подкрепление (награда) усиливает вероятность этой реакции, а негативное (наказание), наоборот, снижает.
Перечисленные выше процессы крайне разнородны по своим нейрофизиологическим механизмам и объединяются в систему имплицитной памяти исходя скорее из их функциональной схожести, а не из-за единого биологического субстрата. Тем не менее, можно выделить некие основные структуры головного мозга, необходимые соответственно для имплицитной и для эксплицитной памяти. Эти структуры указаны на рис. 2. В механизмы имплицитной памяти вовлечены неокортекс, стриатум, миндалина, мозжечок, а в простейших случаях – проходящие через спинной мозг рефлекторные пути сами по себе. Для функционирования эксплицитной памяти необходимы медиальные височные доли, гиппокамп, а также различные отделы коры.

Principles of Neural Science, Fifth Edition, 2012
а. Структуры, необходимые для функционирования различных типов эксплицитной памяти. Striatum – полосатое тело, Neocortex – неокортекс, Amygdala – миндалевидное тело, Cerebellum – мозжечок, Reflex pathways – рефлекторные пути. Структуры, необходимые для функционирования эксплицитной памяти. Medial temporal lobe – медиальные височные доли, Hippocampus – гиппокамп.
Два вида эксплицитной памяти – семантическая и эпизодическая – в большей степени сходны между собой нейрофизиологически, однако и между ними имеются различия. В настоящий момент семантическая память исследована в меньшем объеме, и дискуссия о конкретных структурах и процессах, ответственных за ее функционирование, все еще ведется. Хотя довольно больший объем исследований свидетельствует о том, что для формирования семантических воспоминаний так же, как и для эпизодических, необходима активность гиппокампа и медиальной височной доли, ряд ученых сомневается в данной концепции. В настоящий момент наиболее изучены механизмы именно долговременной эпизодической памяти, поэтому далее речь пойдет именно о них.
Эпизодическая память не представляет собой какой-то единственный непрерывно протекающий процесс. Она состоит из совокупности сменяющих друг друга процессов, обеспечивающих накопление, эффективное хранение и использование имеющихся воспоминаний. Схематично это показано на рис. 3.

Рис.3. Схема процессов эксплицитной эпизодической памяти
Для начала необходимо разобрать, как же происходит кодирование воспоминаний, то есть, в какой форме чувственный опыт сохраняется для последующего использования. Каждое событие в жизни, безусловно, связано с восприятием некоего набора объектов, расположенных в определенном порядке и обладающих чувственными характеристиками. Представим на минуту, что у нас сегодня День рождения, и наши друзья принесли торт, зажгли свечи и поздравляют нас, поя песенку «Happy birthday to you». Мы в этом момент наблюдаем целую сцену, видим, где расположен торт, как стоят наши друзья, слышим, как они поют, чувствуем запах дыма от свечей. Восприятие этой сцены связано с определенным паттерном активности зон коры, ответственных за восприятие и обработку стимулов от органов чувств. Однако спустя некоторое время мы наблюдаем уже другую сцену, наши органы чувств воспринимают уже совсем другие объекты, и активность сенсорных зон мозга совсем иная. К счастью, в головном мозге есть особая структура, позволяющая «записывать» паттерн активности отвечающих за обработку входящей сенсорной информации при восприятии той или иной сцены. Этой структурой является гиппокамп – относительно небольшая парная структура, расположенная в глубине каждой медиальной височной доли.
Значение гиппокампа в процессе запоминания новой информации было наиболее ярко продемонстрировано американскими исследователями Scolville и Milner, описавшими в 1957-м году клинический случай пациента H.M. Этот пациент страдал от тяжелейшей эпилепсии, в связи с чем было принято решение тотально удалить оба гиппокампа. В результате этой операции у H.M. была полностью потеряна способность фиксировать новые воспоминания при сохранении до определенной степени возможности воспроизводить старые, то есть, развилась изолированная антероградная амнезия. Интересно, что, хотя H.M. больше не мог запоминать какие-то новые факты или эпизоды из текущей жизни, его имплицитная память осталась интактной. В ходе множества экспериментально-психологических и когнитивных тестов было показано, что H.M. способен к определенного рода обучению и освоению новых процедурных навыков, хотя никогда и не сможет запомнить, когда и как он им обучался.
Так как же в относительно небольшом гиппокампе способна умещаться информация о нашем восприятии во множестве временных точек. Вернемся к нашему Дню рождения. Активные в данный момент нейроны, отвечающие за восприятие зрительной (вид торта, свечей, друзей), слуховой (песня) и иных видов сенсорной информации, а также пространственная информация (взаиморасположение друзей, торта и всяких разных вещей относительно нас), объединенные вместе в одну связную картину через ассоциативные зоны коры, формируют так называемые «энграммы», то есть, некий «след» восприятия. Соответственно, пути, которые связывают эти зоны с гиппокампом (антероградные) активирующиеся в одно время, постепенно конвергируют и заканчиваются на гораздо меньшей по численности группе нейронов в гиппокампе (эти нейроны называют «энграммными»). Эти нейроны, в свою очередь, имеют отростки, формирующие ретроградные пути, то есть, идущие от гиппокампа обратно к тем самым сенсорным и ассоциативным зонам коры.
Таким образом, при возбуждении этой небольшой группки нейронов (в некоторых случаях достаточно даже активации одного единственного нейрона), активируются гораздо более обширные зоны коры головного мозга, «симулирующие» восприятие определенной имевшей когда-то место сцены, так как паттерн возбуждения этих зон будет практически аналогичен таковому при реальном восприятии, пусть даже это возбуждение и будет несколько слабее. Эта чувственная «симуляция» и будет представлять собой эпизодическое воспоминание.
При этом, чтобы вызвать активность энграммных нейронов, то есть, вызвать воспоминание, не нужно полностью повторить активность, идущую по антероградным путям, иначе мы бы смогли вспоминать что-либо, только увидев абсолютно идентичную картину. То есть, мы бы вспоминали о своем предыдущем дне рождения, только если наши друзья в этот год поздравляли бы нас абсолютно точно так же. Для активации энграммных нейронов гиппокампа и последующей активации сенсорной и ассоциативной коры достаточно лишь частичного совпадения стимулов. Даже запах свечей сам по себе, увиденный торт в магазине, услышанная краем уха по радио поздравительная песня может вызвать у нас развернутое воспоминание о том самом прошлом дне рождения и наших друзьях. И, конечно, вспоминать что-то можно и в полное отсутствие каких-либо ассоциативно связанных стимулов, лишь по собственному желанию. Это возможно благодаря наличию связей между гиппокампом и лобными отделами коры, отвечающими за произвольные действия и за сознательный контроль физической и психической деятельности вообще.
Кроме того, с течением времени, если воспоминание не теряет своей актуальности и сохраняется надолго, постепенно усиливаются связи между самими сенсорными и ассоциативными отделами коры, а связи с гиппокампом, наоборот, ослабевают. В результате этого для воспроизведения полноценного эпизодического воспоминания не нужна будет активация энграммных клеток гиппокампа, а достаточно будет возбудить лишь один участок коры, чтобы активировались все связанные с ним в рамках данного конкретного воспоминания (рис.4.).
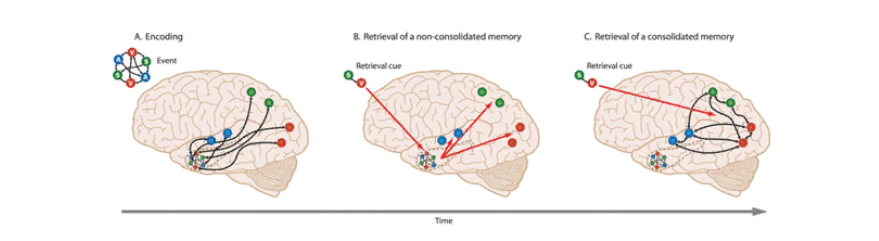
Principles of Cognitive Neuroscience, 2nd ed. 2013.
Рис.4. Кодирование и воспроизведение воспоминаний. А. Кодирование; В. Воспроизведение неконсолидированного воспоминания; кодирование консолидированного воспоминания.
Очевидно, что для поддержания этого процесса необходимо облегчить передачу информации между нейронами, «ответственными» за одно воспоминание так, чтобы лишь малая часть первоначальной импульсации приводила к той же обширной активности. Это «облегчение» и его поддержание и составляет суть процесса консолидации памяти. В его основе лежит феномен долговременной потенциации синаптической передачи (long-term potentiation, LTP), заключающийся в функциональных и структурных изменениях синапса, способствующих большей деполяризации постсинаптической мембраны в ответ на ту же или меньшую деполяризацию пресинаптической мембраны. Традиционно LTP разделяют на два этапа: раннюю LTP и позднюю LTP (см рис. 5)
Большинство нейронов, участвующих в процессах консолидации памяти, глутаматергические, а постсинаптическая мембрана синапса сожержит NMDA-рецепторы, которые при связывании с глутаматом открывают ионный канал, через который в клетку проникают ионы натрия и кальция. Этот внутриклеточный кальций затем связывается с регуляторной молекулой кальмодулином и запускает каскад реакций, приводящих к встраиванию уже созданных клеткой, но до этого находившихся в везикулах, NMDA-рецепторов в мембрану, а также к синтезированию определенных сигнальных молекул, выбрасываемых обратно в синаптическую щель и приводящих к большему выбросу глутамата. Таким образом, происходит достаточно быстрое «облегчение» синаптической передачи. При этом происходят также изменения цитоскелета этого принимающего сигнал дендрита, что будет важно для следующего этапа – поздней LTP. Однако, как было показано выше, этап ранней потенциации связан скорее с функциональными изменениями внутри нейрона, в ходе нее не происходит синтеза новых белковых молекул и каких-то качественных изменений в синапсе. Поэтому эффект от ранней потенциации недолговечен и длится не больше нескольких часов.
На данном этапе заканчивается консолидация не самых нужных для нас воспоминаний. Действительно, мы еще можем вспомнить, если постараемся, внешний вид ничем не примечательных людей, с которыми мы ехали полчаса назад в метро, но на следующий день у нас это уже, скорее всего, не получится. Однако, если те же соседи в метро были чем-то крайне примечательны, и мы целенаправленно и долго на них смотрели, то воспоминание о них, скорее всего, сохранится у нас на гораздо больший срок, что будет возможно благодаря включившимся механизмам поздней потенциации синаптических связей. Более сильный или часто повторяющийся стимул приводит к большему и более частому возбуждению пресинаптической мембраны, что приводит к большему выделению глутамата, большей активации NMDA-рецепторов и, следовательно, большей концентрации кальция внутри нейрона. Если кальция в этом нейроне достаточно много, то помимо связывания с кальмодулином, он также активирует аденилатциклазу, что, спустя несколько этапов, приведет к активации CREB (cAMP-responsive element binding protein). Фосфорилированный CREB затем связывается с CRE-последовательностями определенных генов и активирует их транскрипцию. Это приводит к долговременным клеточным изменениям.
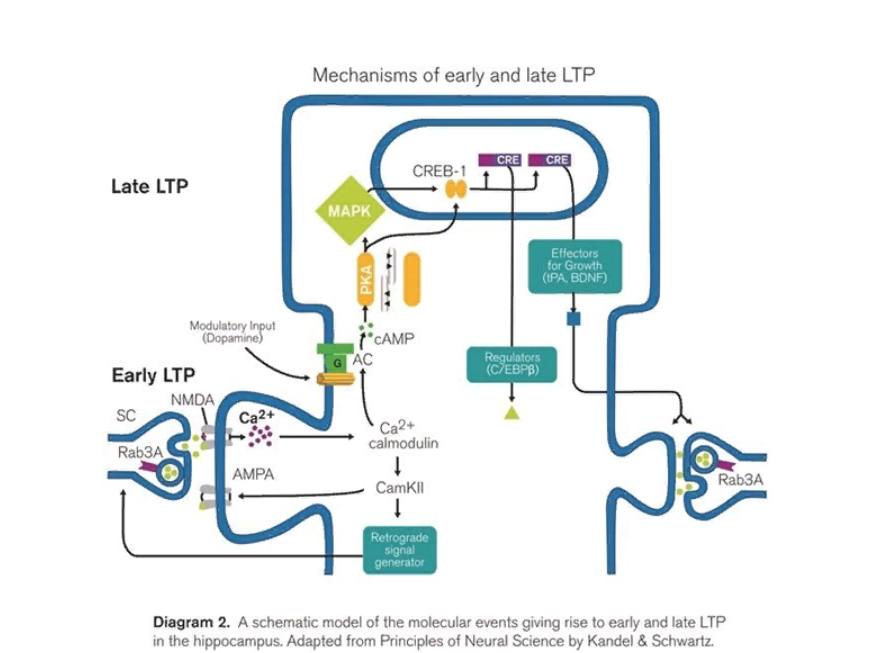
Рис. 5. Биохимические механизмы ранней и поздней потенциации.
Во-первых, продукты этого синтеза закрепляют изменения, произошедшие на раннем этапе LTP, а, во-вторых, они не просто усиливают передачу по одному синапсу, но и увеличивают само число синаптических связей между этими двумя нейронами, что гораздо сильнее и на неопределенно долгий срок упрощает передачу возбуждения от одной клетки к другой и, следовательно, закрепляет воспоминания. При этом, эти изменения специфичны именно для синапса с конкретным нейроном благодаря изменившейся в ходе ранней LTP цитоархитектонике внутри необходимого дендрита. Таким образом, именно эти, на первый взгляд, достаточно простые электро- и биохимические процессы в результате приводят к тому, что мы можем без особых усилий заново «переживать» свой прошлый опыт даже спустя многие десятилетия.
Однако единожды консолидированное воспоминание крайне редко статично хранится в памяти. При каждом осознанном и неосознанном воспоминании, то есть, при каждом новом прохождении импульса по этим «облегченным» путям, они сами становятся в какой-то мере хрупкими и пластичными. Для того, чтобы вернуть эту систему обратно в стабильное состояние, существуют процессы так называемой реконсолидации воспоминаний. Реконсолидация по своим биохимическим принципам во многом сходна с процессами консолидации, однако при ней возможны системные, сетевые, нейронные изменения, в результате чего воспоминания могут усилиться, «обрасти» новыми деталями и аспектами, или, если они уже более не так актуальны, распасться и постепенно исчезнуть.
Процессы распада и исчезновения ненужных воспоминаний у здоровых людей, к сожалению, исследованы в гораздо меньшем объеме Хотя в силу большой, но ограниченной информационной емкости мозга, процессы удаления воспоминаний, определения их актуальности и потребности в сохранении, безусловно не менее важны и интересны, чем все остальные процессы, связанные с памятью. Исследователи предполагают существование множества различных механизмов, отвечающих за дезинтеграцию энграмм и распад воспоминаний – от простого биохимического регресса синаптических изменений до активных процессов, связанных со специальным удалением травмирующей, противоречащей новому опыту или более не нужной информации. Интересными представляются исследования, проведенные на мушках-дрозофилах. Оказалось, что у данных насекомых имеются так называемы «клетки забывания», которые способны посредством дофаминергической передачи и активацию сигнального белка Rac вызывать каскад реакций, разрушающий интеграцию нейронов в энграммы и, соответственно, «стирающих» воспоминание. По-видимому, подобные нейроны активируются при достаточно долгом отсутствии специфических стимулов, вызывающих данное воспоминание, то есть, при его дезактуализации.
Подытожив, хочется отметить, что в данной статье крупными мазками были описаны лишь основные механизмы, связанные с функционированием лишь одного вида памяти – эксплицитной эпизодической. Память представляет из себя невероятно сложный и обширный предмет исследования, разбираться в котором мы только начинаем. Однако в настоящий момент огромное количество исследовательских групп по всему миру углубляет наши знания о памяти. И, скорее всего, в самое ближайшее время именно в этой области стоит ждать громких открытий, детально объясняющих, как же все-таки на самом деле работает мозг, наш самый таинственный орган.
Автор текста: Кибитов А.А.
Список литературы:
- Б. Баарс, Н. Гейдж. Мозг, познание, разум. Введение в когнитивные нейронауки: в 2-х т. / ред. В.В. Шульговского.– 2-ое изд., испр.– Москва: «Лаборатория знаний, 2016 г.
- Kandel ER, Schwartz JH et al. Principles of Neural Science, Fifth edition / McGraw-Hill Education, 2012.
- Schlichting ML & Preston AR. Memory integration: neural mechanisms and implications for behavior. Current Opinion in Behavioral Sciences, 2015, 1, 1–8. doi:10.1016/j.cobeha.2014.07.005
- Purves D, Cabeza R. et al. Principles of Cognitive Neuroscience, 2nd edition. / Sinauer Associates, Inc, 2013
- Frankland PW, Bontempi B. The organization of recent and remote memories. Nat Rev Neurosci 2005;6:119–30.
- Raslau FD, Klein AP, Ulmer JL et al. Memory Part1: Overview. Am J Neuroradiol, 2014, 35(11).
- Plaff DW. Neuroscience in the 21st Century. From Basic to Clinical. 2nd edition. / Springer, 2016
- Puzzo D, Fiorito J, Purgatorio R et al. Genes, Environment and Alzheimer’s Disease: Ch. 1 –Molecular Mechanisms of Learning and Memory / Academic Press, 2016, P.1-27
- Shuai Y, Lu B, Hu Y et al. Forgetting is regulated through Rac activity in Drosophila. Cell. 2010 Feb 19;140(4):579-89. doi: 10.1016/j.cell.2009.12.044.