
Нейрогенез – это процесс образования нейронов. Функционирует он примерно также, как и образование кровяных тел: есть пул клеток предшественников нейронов, происходит их пролиферация, далее происходит их развитие и дифференцировка, а кончается все это образованием нового нейрона, интегрированного в нейрональную сеть. Понятное дело, что этот процесс наиболее активен во время пренатального развития, но все же некоторая его активность наблюдается и у взрослых. В данной статье речь пойдет именно про нейрогенез у взрослых, эмбрионального же развития касаться в основном не будем.
Нейрогенез у взрослых организмов был обнаружен сравнительно недавно (только в последние десятилетия), в основном благодаря развитию иммуногистохимических методов и конфокальной микроскопии. Сначала выявили постнатальный нейрогенез у птиц, затем у млекопитающих и только потом уже у человека [1].
Методы исследования нейрогенеза
Иммуногистохимическое исследование – метод микроскопического исследования тканей, обеспечивающий наиболее специфическое выявление в них искомых веществ и основанный на обработке срезов маркированными специфическими антителами к выявляемому веществу, которое в данной ситуации служит антигеном. Метод бывает прямой и непрямой. Прямой основан на реакции специфического связывания марированных антител непосредственно с антигеном. При непрямом сначала с антигеном связываются антитела, а уже потом с антителами марикрованные антитела.
Конфокальная микроскопия – разновидность световой оптической микроскопии, обладающей значительным контрастом и пространственным разрешением по сравнению с классической световой микроскопией. По сути, такой эффект достигается за счет контролируемого ограничения глубины фокуса оптической системы (Схема 1).
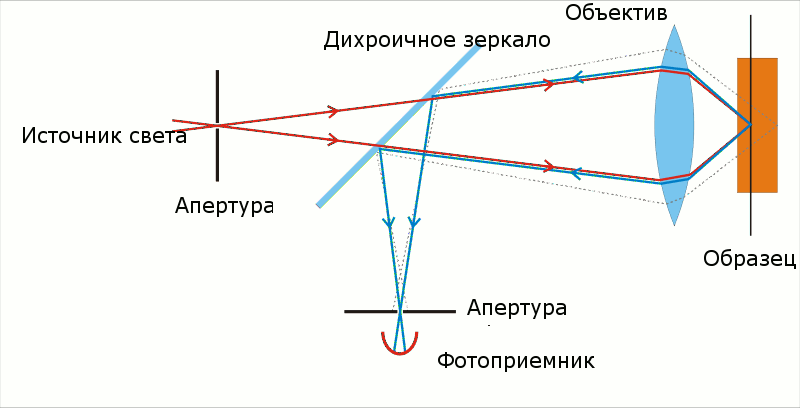
Схема 1
Но что же все искали в клетках, участвующих в нейрогенезе? А искать нужно белок даблкортин (DCX). Этот белок ассоциирован с микротрубочками и экспрессируется практически всеми незрелыми нейронами. Нейрональные клетки-предшественники начинают производить DCX вскоре после входа в клеточный цикл с затуханием экспрессии через 2-3 недели, ко времени окончательного превращения в развитые нейроны.
Имеется также еще один способ изучения нейрогенеза. Заключается он в использовании бромдезоксиуридина (BrdU). Он является синтетическим нуклеозидом, аналогом тимидина, используемым для выявления пролиферирующих клеток в живых тканях, а также для изучения репликации ДНК. Бромдезоксиуридин способен заменять тимидин в процессе репликации ДНК, встраиваясь в новую ДНК. Иммуногистохимическое окрашивание с антителами к бромдезоксиуридину позволяют обнаружить включённый модифицированный нуклеозид, тем самым выявляя пролиферирующие клетки [2,3,4].
Но где же находятся эти пулы стволовых клеток? Есть два основных места: субгранулярная зона зубчатой извилины гиппокампа и субвентрикулярная зона.
Субгранулярная зона зубчатой извилины гиппокампа
Субгранулярная зона зубчатой извилины гиппокампа – область мозга, расположенная между слоем гранулярных клеток и хилусом зубчатой извилины гиппокампа. В субгранулярной зоне происходит постнатальный («взрослый») нейрогенез — образование новых нейронов из полипотентных клеток-предшественников [5].
Субвентрикулярная зона
Субвентрикулярная зона – это область первичной миграции нейро- и глиобластов из вентрикулярной герминативной зоны. Субвентрикулярная зона простирается вдоль большей части латерально расположенной внутренней поверхности боковых желудочков мозга. Наряду с субгранулярнойзонойзубчатой извилины гиппокампа, субвентрикулярная зона является источником новых нейронов на протяжении всей взрослой жизни организма (Рис. 1).
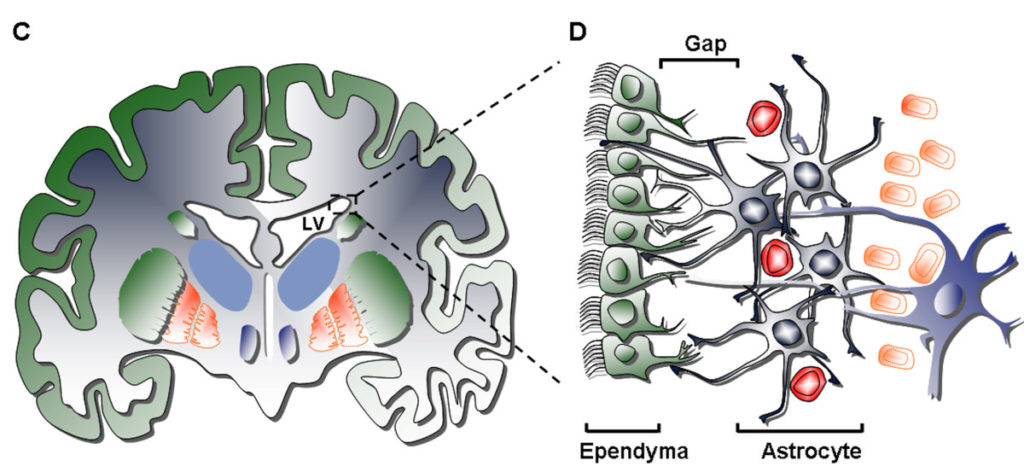
Рис. 1
Выделяется 4 типа клеток субвентрикулярной зоны:
1) Реснитчатые эпендимоциты типа E
2) Пролиферирующие нейробласты типа А
3) Медленно пролиферирующие клетки типа B
4)Активно пролиферирующие клетки типа С
Реснитчатые эпендимоциты типа Е обращены в полость желудочка и стимулируют циркуляцию цереброспинальной жидкости. Пролиферирующие нейробласты типа А объединяются в цепочки и мигрируют по направлению в обонятельной луковице. Медленно пролиферирующие клетки типа B образуют глиальные трубки, внутри которых происходит миграция нейробластов типа А. Активно пролифирующие клетки типа С образуют скопления между цепочками клеток А [6].
Ростральный миграционный тракт
Ростральный миграционный тракт – это путь по которому нейробласты мигрируют из субвентрикулярной зоны в обонятельную луковицу. Зарождение новых клеток в субвентрикулярной зоне и их миграция по ростральному миграционному потоку происходит на всем протяжении взрослой жизни организма. Группы нейробластов мигрируют цепочками, продвигаясь по глиальным трубкам, образованным астроцитарными клетками и их отростками.Лишь достигнув середины обонятельной луковицы, цепочки новорожденных нейронов распадаются и клетки начинают радиальную миграцию. Так они достигают верхних клеточных слоев, где происходит их окончательная дифференциация. Рассеивание цепочек нейробластов инициируется протеинами рилином и тенасцином, а сам процесс радиальной миграции зависит от наличия тенасцина-R. Большое количество новых нейронов отмирает вскоре после окончания миграции. В долгосрочной перспективе, около 50 % оставшихся клеток также отмирают, даже после успешного приживления в гранулярном и перигломерулярном слоях и установления связей с другими клетками [7].
Функциональное значение нейрогенеза – это все еще дискутабельный вопрос, вдруг он является просто сбоем механизма эмбрионального развития? Но все же считается (и волне обосновано), что нейрогенез способствует пластичности биологических нейронных сетей. Например, молодые гранулярные клетки в зубчатой извилине имеют более низкий порог долговременной потенциации, чем старые клетки [8].
Регуляция нейрогенеза
Но что же регулирует процесс нейрогенеза у взрослых? Почему у нас нет бесконечно делящихся нейронов и наш мозг не разрастается до того, что перекрывает все пути циркуляции ликвора? Факторы ответственные за этот процесс весьма многочислены, вот лишь некоторые из них: гормоны, факторы роста, различные нейротрансмиттеры, цитокины, электрофизиологическая активность. Ниже мы рассмотрим эти процессы чуть болеее подробно.
С помощью различных манипуляций с дофамином учёным удалось влиять на интенсивность нейрогенеза. Что касается участия серотонина в нейрогенезе взрослого мозга, установлено его потенцирующее влияние на образование и трансформацию нейробластов в субвентрикулярной зоне гиппокампа. Активация 5-НТ2С рецепторов в регионе сосудистого сплетения вдвое увеличивает пролиферацию прогениторов в субвентрикулярной зоне, тогда как блокада 5-НТ2А рецепторов ведет к аналогичному снижению числа пролиферирующих клеток в субгранулярной зоне гиппокампа. Более того, систематическое введение ингибиторов синтазы оксида азота увеличивает пролиферацию в субвентрикулярной зоне, но не в зубчатой извилине гиппокампа. ГАМК также усиливает нейрогенез, причём некоторые из его эффектов реализуются через белок CREB. Действие же глутама двояко (он может как снижать, так и повышать пролиферацию и дифференцировку) и зависит от рецептора, с которым он связывается. Наконец можно обрадовать и тех, кто ищет паралели с болезнью Альцгеймера – ацетилхолин. Нарушение холинэергической иннервации снижает число образующихся нейронов, а усиление – наоборот.
РАСАР (Pituitary Adenylate Cyclase Activating Polypeptide – «полипептид, активирующий аденилатциклазу гипофиза»). Данный пептид реализует свои эффекты через РАС1R рецептор. Стволовые клетки субвентрикулярной зоны при внесении агониста рецептора РАС1R пролиферировались in vitro, тогда как селективный антагонист РАСАР тормозил этот процесс.
Также не следует забывать о нейротрофинах. Нейротрофины – семейство крупных полипептидов, которые регулируют выживание, развитие и согласованную функцию нейронов. Секретируемые нейрональными и васкулярными клетками нейротрофины выполняют сигнальную миссию в большом спектре физиологических процессов. Эффекты нейротрофинов осуществляются при их взаимодействии с тирозинкиназными рецепторами. Принципиально и то, что после соединения рецептора с лигандом запускается каскад сигнальных реакций, специфичных для этого контакта. Нейротрофины активируют тирозинкиназные рецепторы семейства Trk – trk-A, trk-B, trk-C.
Поскольку нейральные стволовые клетки способны дифференцироваться в различные типы – нейроны, олигодендроциты и глиальные клетки, вектор их трансформации, по-видимому, определяется специфическими путями сигнального контроля. Регуляция таких каскадных реакций зависит от ростовых и нейротрофических факторов. Выделяют основные соединения, которые играют роль в трансформации НСК: (1) нейротрофический фактор мозга (BDNF); (2) тромбоцитарный и эпидермальный факторы роста (PDGF/EGF); (3) трансформирующий ростовой фактор (TGF-beta1) и костный морфогенный белок (BMP); (4) группа, включающая интерлейкин-6 (IL-6), ингибирующий фактор лейкемии (LIF), реснитчатый нейротрофический фактор (CNTF). Эти вещества рассматриваются как лиганды, которые после соединения с рецептором индуцируют специфические сигнальные процессы. В результате экспрессируется активность внутриклеточных мессенджеров и далее – селективная активация транскрипторных факторов, в соответствии с функциональным назначением сигнала.
Реализация любых эффектов нейротрофинов осуществляется при участии тирозинкиназных рецепторов с последующим включением трансдукторных реакций. Активация тирозинкиназных рецепторов стимулирует многовариантный каскад реакций, ведущих к образованию продуктов с последовательной экспрессией (фосфорилированием) новых белков. Результатом этих процессов оказывается рилизинг Са++ в цитоплазме нейрона, торможение апоптоза и стимулирование выживания клеток. Далее, результатом трансдукторных реакций является перенос сигнала в ядро нейрона и стимуляция транскрипторных процессов, реализуемых на уровне ДНК, осуществляющих синтез продуктов, необходимых для функции клеток.
Активация тирозинкиназных рецепторов ведет к инициации молекулярных сигналов, нацеленных на экспрессию генов, кодирующих соответствующие функциональные белки. Активация TrKs ведет к активации PLC-γ (фосфолипаза С-γ), PI3K (фосфатидилинозитол-3- киназа) и PIP2 (фосфатинозитол 3,5–бифосфат). Путь сигнальной регуляции, указанный в правой части рисунка, предусматривает после активации PLC-γ образование DAG (диацилглицерол) и IP3 (инозитолтрифосфат). Молекулы, рассматриваемые как вторичные посредники, стимулируют активность РКС-δ (протеинкиназа-δ) и увеличение внутриклеточного Са2+; эти процессы являются ключевыми в активации кальмодулина (СаМ), образования и дифференцировки нейронов. Еще один путь включает активацию молекул семейства Ras (небольшие G-белки гуанитидилтрифосфатазы), далее белки Raf-киназы, промотирующие, в свою очередь, фосфорилирование и активацию системы MAPK/ERK (MAPKs – митоген-активированные белки и ERK1/2 – внеклеточные сигнал-регулируемые киназы). Эти фосфорилированные белки транслоцируются далее в клеточное ядро, где они участвуют в активации транскрипторных факторов CREB, NF-kB, c-JIN (и др.), регулируя экспрессию генов, промотирующих формирование нейронов. Например, связывание нейротрофина NGF с рецептором TrKA в терминалях аксонов стимулирует рост последних как результат «прозванивания» цепочки ERK1/2- CREB. На культуре симпатических нейронов было установлено, что торможение сигнального пути Ras-PI3K-Akt снижает выживание трансформирующихся клеток после апоптоза. MAPK-ERK1/2 участвуют в нейрогенезе, контролируя селекцию клеток путем индукции генов выживания и торможением проапоптических белков. Ras также подавляет апоптоз через экспрессию PI3K-Akt, которые блокируют проапоптические белки.
Подготовил: Коровин А.С.
Источники:
1 – Ming G. L., Song H. (2011). «Adult neurogenesis in the mammalian brain: significant answers and significant questions.». Neuron 70 (4): 687—702. DOI:10.1016/j.neuron.2011.05.001.
2 – Wilson G. D. et al. Measurement of cell kinetics in human tumours in vivo using bromodeoxyuridine incorporation and flow cytometry //British journal of cancer. — 1988.
3 – Gratzner H. G. Monoclonal antibody to 5-bromo-and 5-iododeoxyuridine: a new reagent for detection of DNA replication //Science. — 1982
4 – Ikushima T., Wolff S. Sister chromatid exchanges induced by light flashes to 5-bromodeoxyuridine-and 5-iododeoxyuridine substituted Chinese hamster chromosomes //Experimental cell research.
5 – Oomen C. A., Girardi C. E., Cahyadi R., Verbeek E. C., Krugers H., Joëls M., Lucassen P. J. Opposite effects of early maternal deprivation on neurogenesis in male versus female rats. (англ.) // Public Library of Science ONE. — 2009. — Vol. 4, no. 1. — P. e3675. — DOI:10.1371/journal.pone.0003675.
6 – Abrous DN, Koehl M, Le Moal M. (2005) Adult neurogenesis: from precursors to network and physiology.
7 – Roy NS, Wang S, Jiang L, et al (March 2000).«[10.1038/73119 In vitro neurogenesis by progenitor cells isolated from the adult human hippocampus]». Nat. Med. 6 (3): 271–7. DOI:10.1038/73119.
8 – Lledo PM (2006). «Adult neurogenesis and functional plasticity in neuronal circuits.». NatRevNeurosci 7 (3): 179-93.
Добавить комментарий