
В обработку визуальных сигналов вовлечено большое количество структур мозга, взаимосвязи которых многочисленны и до конца не изучены. Информация об анализе визуальных стимулов, которой мы обладаем на данный момент, по крупицам собрана из огромного количества отдельных исследований. Каждое исследование предоставляет результаты одного или серии экспериментов, а их сумма позволяет составить общее впечатление о некоторых аспектах работы головного мозга, доказать или опровергнуть выдвигаемые гипотезы.
Визуальная система часто изучается в ходе фундаментальных исследований в области нейронаук по ряду причин. Во-первых, она связана со зрением — основным каналом получения информации из окружающего мира, но при этом она также узкоспециализирована, что позволяет разрабатывать разнообразную методологию исследований. Во-вторых, область зрительной коры удобна для изучения на обезьянах с использованием инвазивных методов регистрации активности мозга в виду своего расположения; в экспериментах с участием людей успешно применяются неинвазивные методы. Кроме того, спектр вопросов, которые представляется возможным прояснить в ходе исследований, достаточно широк: аспекты осознанного/неосознанного восприятия, природа воображения, обработка и фильтрация визуальной информации, распределение внимания, повреждения мозга и связанные с ними расстройства и др. В данной статье мы сосредоточимся в основном на первичной зрительной коре, оговорим предшествующий ей путь нервных сигналов и некоторые общие свойства зрительной коры.
Визуальная система
Когда мы видим изображение, ганглионарные клетки сетчатки генерируют нервные импульсы и передают их в латеральное (оно же наружное) коленчатое тело (ЛКТ), которое расположено в таламусе. Оно состоит из шести слоев, первые два из них представлены магноцеллюлярными клетками, остальные четыре — парвоцеллюлярными. Магноцеллюлярные клетки передают информацию об изображениях с низкой контрастностью, движущихся объектах, они не восприимчивы к цвету, их сигналы быстрые и кратковременные, они дают представление о воспринимаемой информации в целом, то есть, быстро и схематично, в низком разрешении. Парвоцеллюлярные клетки чувствительны к цвету и лучше воспринимают высококонтрастные изображения, они передают более медленные и длительные сигналы, что позволяет получить более детальную, хотя и медленную информацию.
Через латеральное коленчатое тело сигналы передаются далее в затылочные доли обоих полушарий, которые ответственны за обработку зрительных стимулов. Первая кортикальная область, куда попадают эти сигналы — первичная зрительная кора (V1). V1 расположена в заднем полюсе затылочных долей, это самая древняя и простая из кортикальных зон, однако, наиболее изученная. V1 обрабатывает информацию о движущихся и статичных объектах, отвечает за распознавание простых образов (например, геометрических форм).
V1 состоит из шести слоев, наибольшее количество аксонов ЛКТ подходит к IV слою, который разделяется еще на четыре подслоя. Клетки V1 бывают двух видов: простые и сложные. Простые клетки встречаются в слоях IV и VI, они реагируют на ориентацию (угол), расположение (относительно центра визуального поля) и яркость объектов. По строению они имеют возбуждающий центр и тормозящую периферию или наоборот (см. рис.). Их ответ на стимул прямо пропорционален соответствию этого стимула «идеалу». Другими словами, у клетки есть «идеальный» стимул, в ответ на который реакция будет наиболее интенсивна, чем дальше стимул от «идеального», тем менее интенсивна реакция. Сложные клетки находятся в слоях II, III, и V, они также имеют предпочитаемую ориентацию, но не чувствительны к местонахождению и яркости объекта. Сложная клетка совмещает в себе две простые клетки с совпадающей предпочитаемой ориентацией, центр клетки полярен периферийным частям.
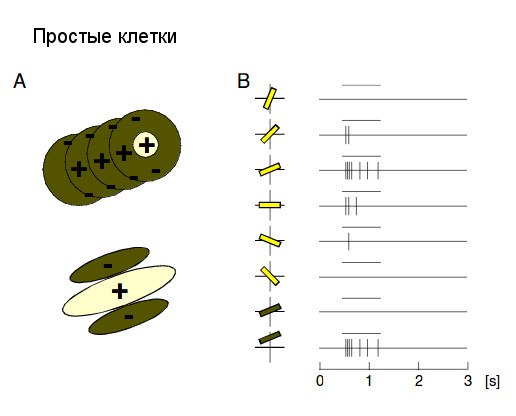

Разница реакций простых и сложных клеток
Условия эксперимента: несколько оптимально ориентированных линий движутся через визуальное поле.
Реакция простых клеток: Клетки реагируют синусоидальными колебаниями мембранного потенциала в соответствии с чередованием черных линий и просветов, проходящих через визуальное поле. Потенциалы действия возникают только в фазе деполяризации.
Реакция сложных клеток: Наблюдается постоянная деполяризация, потенциалы действия выглядят беспорядочными.

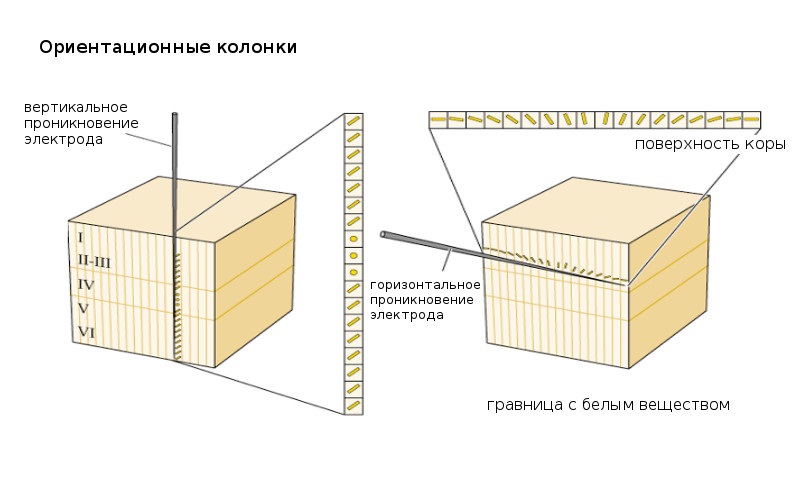
Ice Cube Model
Эта гипотетическая кубическая модель придумана для пояснения устройства клеток первичной визуальной коры, а именно – как устроены предпочитаемые ориентации и, соответственно, реакции нейронов V1. Так, V1 можно условно поделить на кубы 2 ммˆ3, каждый из которых получает сигналы от обоих глаз. Клетки с одинаковыми ориентационными предпочтениями формируют горизонтальные колонки, при этом соседние вертикальные колонки имеют слегка отличающиеся ориентационные предпочтения.
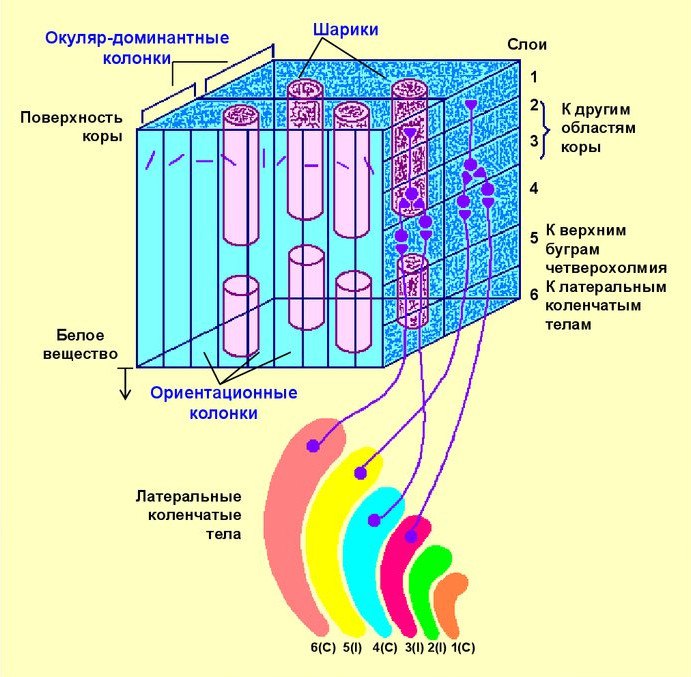
Чувствительные к цветам клетки также собраны в столбцы (также их называют каплями, гиперколонками, шариками) 0,5 мм в диаметре в зонах соответствующих превалирующих глаз (картинка с цилиндрами). Каждый такой столбец содержит реагирующие либо на красно-зеленый, либо на сине-желтый контрасты.
Оптическая репрезентация карты зрительной коры у млекопитающих (кошки)
Суть эксперимента: Данные регистрируются инвазивным способом. В черепной кости делается отверстие в необходимой зоне (в данном случае V1), кора подсвечивается, на нее направляется линза и камера, которая позволяет регистрировать изменение кровяного потока. Данные регистрируются до и после предъявления животному стимула (линии с определенной ориентацией), две картинки сравниваются для выявления наиболее активных в момент демонстрации стимула зон. Эксперимент повторяется много раз со стимулами разной ориентации, для каждой из них берется сумма значений.
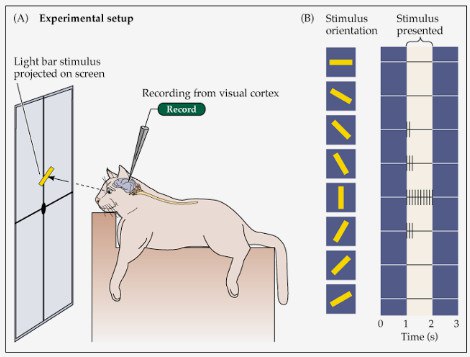
Подписи к картинке: (А) Организация эксперимента: – экран, на котором показана светлая полоска; – регистрация сигналов со зрительной коры. (В) – ориентация презентуемых стимулов; – реакция на стимулы; – время (секунды).
Затем каждая ориентация кодируется определённым цветом для построения карты, где цвета накладываются друг на друга и отображают скопления нейронов с одинаковыми ориентационными предпочтениями, кроме того, соседние цветовые сегменты карты имеют похожие предпочтения. На пересечениях цветовых сегментов ориентационное предпочтение быстро меняется упорядоченным образом, т.е. в этих областях происходят отклики на стимулы с разной ориентацией. Однако данный эксперимент измеряет активацию нейронов только косвенным образом. Вывод можно сделать следующий: организация кортикальных нейронов в аспекте ориентационных предпочтений несколько сложнее, нежели в кубической модели.
Составление ориентационных карт:
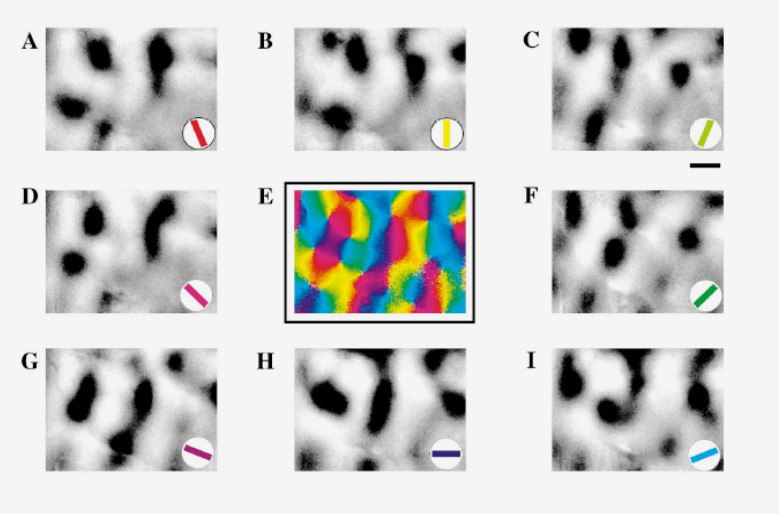
Ориентация и зрение

Подписи к картинке: (А) ориентационные предпочтения; (В) окулярная доминантность – пересечения – пики доминантности; (C) пики пересечений и окулярной доминантности на карте доминантности; (D) бинарная карта окулярной доминантности с пересечениями
Свойства образования топографической карты в зрительной системе
В ретинотопических картах соседние клетки сетчатки представлены соседними клетками V1, такая карта демонстрирует изоморфизм и непрерывное отображение. Также как в других полушарных структурах мозга, репрезентация левого визуального поле отражается в правой части зрительной коры и наоборот. Также ввиду большего количества рецепторов в центре сетчатки, он шире представлен в зрительной коре, нежели периферия. В топографической карте отображаются: ориентационные предпочтения, доминирующий глаз, пространственное разрешение.
Слепое зрение
Слепое зрение — возможность видеть и распознавать объекты, будучи неосведомленным об этом. Феномен проявляется в некоторых случаях повреждения зрительной коры и говорит о том, что видеть и быть осведомленным — разные мозговые функции.
Условия эксперимента, доказывающего феномен слепого зрения: субъекту предъявляются стимулы, которые движутся либо в одну, либо в другую сторону. И хотя субъект утверждает, что не видит их, при просьбе его «угадать» в какую сторону двигался объект, то правильные ответы статистически значительно превышали случайную вероятность. Из этого эксперимента можно заключить, что сетчатка может иметь путь передачи визуальной информации помимо латерального коленчатого тела, и эта информация каким-то образом анализируется мозгом.
Подготовила: Алмазова Т.А.
Источники:
H. Hubel, T. N. Wiesel. Receptive fields of single neurones in the cat’s striate cortex, – J Physiol. 1959 Oct; 148(3): 574–591.
Carandini, D. Ferster Membrane. Potential and Firing Rate in Cat Primary Visual Cortex, – Journal of Neuroscience, 1 January 2000, 20 (1) 470-484.
G. Matthews. Neurobiology: Molecules, Cells and Systems, – Blackwell Science, 1998.
К. Ю. М. Смит. Биология сенсорных систем, – М.: БИНОМ, 2013.