
Смещение внимания от отдельных областей к связям между ними – одна из основных тенденций когнитивной нейронауки. Важность связей между «сенсорным» и «моторным» речевыми центрами подчеркивалась еще первыми исследователями речи. Наряду с зонами Брока и Вернике основной структурой в классической модели речи считается аркуатный пучок, который их связывает. Взамен этой модели ученые предлагают гораздо более обширную схему, основанную на трактографических исследованиях.
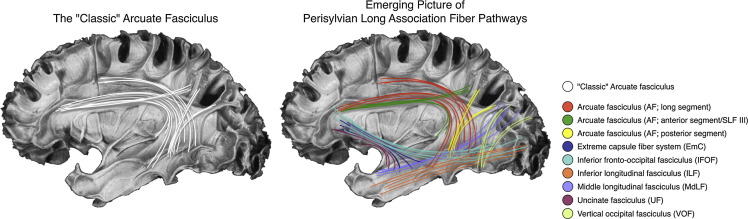

Морфологически выделяют несколько групп проводящих путей, которые так или иначе связывают другие отделы мозга с височной корой:
1) Лобно-височная группа связей включает аркуатный, унцинатный, нижний лобно-затылочный пучки, а также группу волокон самой наружной капсулы.
-аркуатный пучок – идет от покрышечной части лобной коры к задним областям височной.
-унцинатный пучок соединяет орбитальную и латеральную лобную кору с полюсом височной доли, передними отделами височной коры, парагиппокампальной извилиной и миндалиной.
– система волокон самой наружной капсулы (extreme capsule fiber system) соединяет предположительно вентральные и латеральные отделы лобной доли с верхними и средними -височной коры.
– нижний лобно-затылочный пучок соединяет нижние и средние отделы затылочной коры (а также, возможно, медиальные отделы теменной) с проекциями от вентральных отделов височной доли и идет к нижней лобной извилине, медальным и орбитальным отделам лобной коры, полюсу лобной доли.
2) Теменно-височная группа включает
–верхний продольный пучок (SLF), в котором выделяют несколько частей: SLF I, SLF II, SLF III и SLF-tp. Для речевой функции важны SLF II (соединяет угловую извилину с дорсальной премоторной и префронтальной корой), SLF III (соединяет надкраевую извилину с вентральной префрональной корой) и SLF-tp (идет от нижней теменной дольки к задней теменной коре).
– средний продольный пучок соединяет заднее-верхние отделы височной доли с верхними и нижними отделами теменной, а также возможно имеет ответвления к затылочной доле.
3) Затылочно-височная группа
– нижний продольный пучок соединяет вторичные зрительные области затылочной доли со средней и нижней височными извилинами, полюсом височной доли, парагиппокампальной извилиной, гиппокампом и миндалиной.
– вертикальный затылочный пучок имеет отношение к таким аспектам речевой функции, как чтение и письмо. Он соединяет латеральную затылочно-височную извилину и борозду с нижними, и, возможно, также верхними отделами теменной доли.
Перечисленные пути могут быть классифицированы иначе, в зависимости от их функионального значения и принадлежности к дорсальному и вентральному потокам информации. Подробнее концепция двух потоков описана в статье по Нейроанатомии речи; она предполагает, что основные проводящие пути, имеющие отношение к речевой функции организованы в два потока информации, из которых дорсальный необходим для связи с моторной системой, а вентральный – для смысловой обработки информации. Экспериментальные исследования не во всем поддерживают эту теоретическую концепцию.

К дорсальному потоку традиционно относят верхний продольный пучок, особенно SLF III, и аркуатный пучок. Длительное время считалось, что при поражении этой группы связей развивается кондукционная афазия: состояние, при котором пациент пишет и говорит с ошибками, но неплохо понимает речь. В дальнейшем выяснилось, что это не так: этот синдром может развиваться и при корковых поражениях, а к очерченному синдрому кондукционной афазии не приводит даже агенезия SLF и аркуатного пучка.
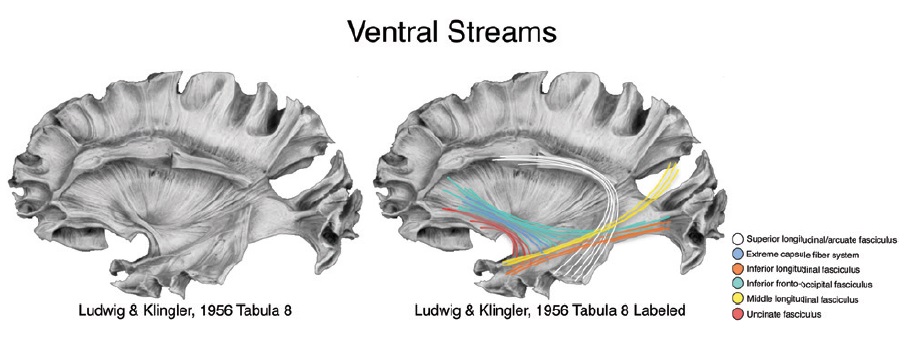
К вентральному потоку относятся волокна самой наружной капсулы, унцинатный пучок, нижний и средний продольные пучки, нижний затылочно-лобный пучок. Многие из перечисленных путей достаточно трудно при исследовании разделить между собой.
Не менее важная функция у путей, которые обеспечивают моторную составляющую речи. Эти связи также можно разделить на несколько групп.
1) собственно нисходящие пути, в первую очередь кортикобульбарные
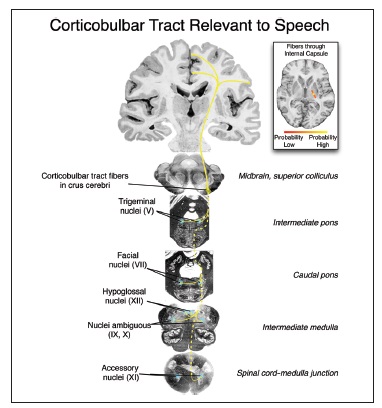
Кортикобульбарные пути обеспечивают связь между ядрами черепно-мозговых нервов и корковым представительством орофарингеальной мускулатуры: вентральными отделами первичной сенсомоторной коры, премоторной коры, и соответствующими соматотопическими отделами добавочной моторной коры. Кроме кортикобульбарных в обеспечении речевоспроизведения участвуют кортикоспинальные пути, которые необходимы для иннервации дыхательной мускулатуры.
2) ассоциативные двигательные пути

Относительно недавно была исследована роль лобного косого пучка. Он соединяет левую дополнительную моторную (SMA) и премоторную (pre-SMA) область с задними отделами нижней лобной извилины. Было показано, что сниженная вербальная беглость у пациентов с первичной прогрессирующей афазией относится к его патологии. Интересно, что он связывает одновременно моторные области (SMA) и отделы, которые относятся к префронтальной коре (pre-SMA), и, возможно, участвует в когнитивном контроле, планировании, подборе последовательности высказывания.
3) корково-подкорковые петли
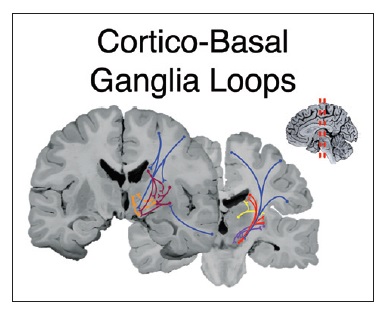
Сложная сеть проводящих путей связывает области коры, участвующие в речевой продукции, с подкорковыми образованиями. Структура этих путей у человека и именно в отношении речевой функции была изучена сравнительно недавно, хотя общее устройство связей коры с субкортикальными ядрами, необходимых для экстрапирамидной регуляции движений, в целом понятно из экспериментов на приматах.
Проекции аксонов первичных и вторичных моторных областей направляются к задней трети стриатума (задние отделы скорлупы и хвостатого ядра). Отсюда проводящие пути идут к наружному и внутреннему бледному шару, субталамическим ядрам и вентролатеральному таламическому ядру. Последнее, в свою очередь, вновь посылает проекции к коре, формируя моторный компонент кортико-стриарной петли. pre-SMA, а также префронтальная кора связаны с передними отделами стриатума, которые, в свою очередь, связываются с внутренней частью бледного шара и вентральными передними ядрами таламуса. Далее волокна вновь идут к префронтальной коре и pre-SMA, замыкая ассоциативную кортико-стриарную петлю, участвующую, помимо организации речи, в когнитивных процессах.
4) корково-мозжечковые связи
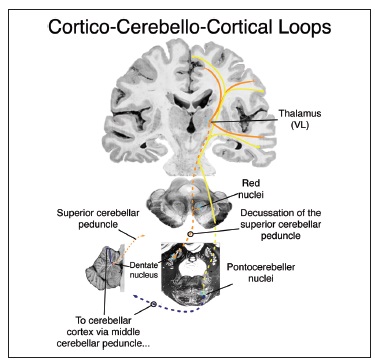
Принципиально не отличается от общей для всех двигательных система кортико-церебеллярно-кортикальная петля, которая связывает ядра мозжечка с собственно моторной и префронтальной корой.
Интересно изучить, как все назаванные области и проводящие пути между ними функционируют совместно, в режиме реального времени. Сети, которые соответствуют активности при выполнении определенного задания, образуют уже не морфологический, а функциональный коннектом. Этот подход к изучению речи принципиально отличается от ранее представленных, которые всего лишь позволяют рассмотреть живой мозг с той же точки зрения, с которой подходили к нему первые исследователи речи.

Так выглядит в визуальном представлении активность мозга в состоянии рассредоточенного внимания (restingstate) и при выполнении различных заданий, как связанных, так и не связанных с речевой функцией. Результаты таких экспериментов показывают, что сложность системы и характер связей между ее элементами резко возрастают и видоизменяются даже при переходе между такими схожими для экспериментатора заданиями, как называние слогов и спонтанная речевая продукция.
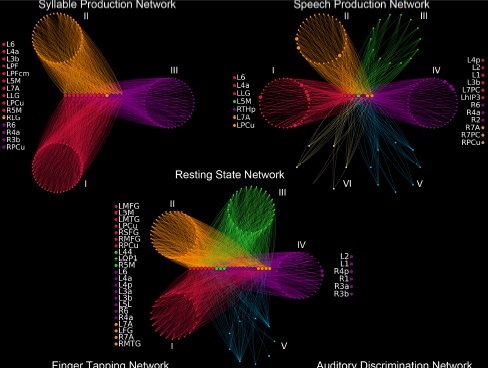
И то, и другое имеет отношение к речевой функции, но функциональный коннектом спонтанной, осмысленной речи оказывается ближе по форме к коннектому состояния рассредоточенного внимания.
Критика исследований речевого коннектома
Имеющиеся данные об областях и проводящих путях основанным в основном именно на таких заданиях, как называние слогов, и других искусственных экспериментальных ситуациях, призванных разделить семантические, синтаксические и другие аспекты речи. Вопрос, правомерно ли экстраполировать результаты таких экспериментов на «натуралистическую речь», специально не изучался. Возможно, это неправильная тактика, которая просто не позволяет нам выявить функциональные связи, актуальные для спонтанной речи. В частности, она приводит к тому, что данные, которые не укладываются в такую «экстраполирующую концепцию», игнорируются или интерпретируются с логическими неточностями.
Например, в эксперименте получают данные об активации области, которых исследователи не ожидали. Вывод делается не о том, что у этой области есть новая функция, а о том, что функция, ранее известная для этой области, необходима для выполнения экспериментальной задачи. Например, по активации гиппокампа делают вывод о том, что задача требует вовлечения механизмов памяти, и только впоследствии исследования подтверждают, что у гиппокамп реагирует на новизну при восприятии вербальных стимулов.
Еще одна возможная ошибка: некая область активируется при решении заданий на различение семантических конструкций, из чего делается вывод, что она специализирована для этой семантической функции. Например, выдвигалось предположение, что зона Брока ответственна за распознавание союзных слов в сложноподчиненных предложениях (как, который и т.п.). Однако последующие эксперименты выявили, что это лишь проявление более общей, базовой функции зоны Брока, которая имеет отношение не к распознаванию семантических конструкций и даже не к языку вообще, а к рабочей памяти: чтобы оперировать такими конструкциями, нужно удерживать в памяти, к чему это союзное слово относится. Принимать в расчет такие факты особенно важно, потому что с точки зрения эволюции речь, скорей всего, формировалась с опорой на предсуществующие когнитивные механизмы, и многое из того, что мы считаем для нее специфичным, может оказаться не «речевым», а «базовым» когнитивным свойством. Отдельные работы дают очень интересные результаты: например, если история преподнесена не в словесном выражении, а в форме последовательных звуков (выстрела, удаляющихся шагов), то при ее восприятии так же задействуются области «высокоуровневой» обработки лингвистической информации (как, например, заднелатеральные височные и нижнее-лобные отделы), как если бы она была изложена в словах.
На данном этапе сложно судить, являются ли «языковыми» такие типичные подходы нашего мозга к обработке информации, как ее сегментирование, формирование представления о типичном для ситуации паттерне действий и основанное на этом паттерне прогнозирование. В частности потому, что иследований, которые рассматривали бы «натуралистическую речь», изучали речь на более высоком, сложном уровне – например, дискурса – критически недостаточно. Именно эти исследования были бы интересны психиатру, потому что на этом уровне, реализуется, по всей видимости, патология мышления.
Автор перевода: Шишковская Т.И.
Источники:
- Tremblay P., Dick A. S. Broca and Wernicke are dead, or moving past the classic model of language neurobiology //Brain and language. – 2016. – Т. 162. – С. 60-71.
- Chang E. F., Raygor K. P., Berger M. S. Contemporary model of language organization: an overview for neurosurgeons //Journal of neurosurgery. – 2015. – Т. 122. – №. 2. – С. 250-261.
- Dick A. S., Bernal B., Tremblay P. The language connectome: new pathways, new concepts //The Neuroscientist. – 2014. – Т. 20. – №. 5. – С. 453-467.
- Fuertinger S., Horwitz B., Simonyan K. The functional connectome of speech control //PLoS Biology. – 2015. – Т. 13. – №. 7.
- Hasson U. et al. Grounding the neurobiology of language in first principles: The necessity of non-language-centric explanations for language comprehension //Cognition. – 2018. – Т. 180. – С. 135-157.