
В последние годы в биологической психиатрии доминирует нейромедиаторная теория. Действительно, актуальность нейромедиаторной теории подтверждается большим числом биологических исследований (содержание нейромедиаторов в ткани мозга, изучение полиморфизмов генов, вовлеченных в обмен нейромедиаторов), а также клинической практикой, ведь успешное применение препаратов, действующих на систему нейротрансмиттеров, предопределяет справедливость теории. Однако есть и альтернативные теории, набирающие популярность. Об одной из таких теорий доложил доктор Ил Хван Ким (Dr. Il Hwan Kim) из университета Дюка в рамках своего семинара в центре наук о здоровье университета штата Теннессии (UTHSC). Его доклад назывался “Изучение нейрональных сетей мозга, относящихся к психиатрическим симптомам” (изобр.[1]).

Изображение 1. Объявление о семинаре (слева), доктор Ил Хван Ким в лаборатории (справа).
Наверняка многие врачи и биологи, да и в целом все, кто изучал физиологию в высших учебных заведениях, слышали про белок актин. Вероятнее всего слово актин сразу ассоциируется со словосочетанием актин-миозиновый комплекс, вспоминаются нервные волокна и дикие для второкурсника теории мышечного сокращения. Кто-то вспомнит про цитоскелет – белковые структуры, которые поддерживают форму клетки такой, какой ей и следует быть. Сразу же из глубин памяти вылезут туманные микрофиламенты, тубулины и кератины. Однако если вы хорошо знакомы с нейроцитологией и нейрогистологией, то вспомните также, что актин играет важную роль в синапсообразовании: небольшие отростки дендритов (дендритные шипики) могут появляться и исчезать, помогая при этом формировать новые синапсы на своей поверхности. Именно благодаря дендритным Шипкам и синаптической динамике нервная система может эффективно обрабатывать и реализовывать информацию и поддерживать свою пластичность в целом.
Формирование дендритных шипиков основано на явлении полимеризации актина, а также ветвлении актиновых структур. Как видно из рисунка [изобр.2], для ветвления актина требуется специальный комплекс белков Arp2/3. Этот комплекс “садится” на материнский актиновый филамент, и на его базе начинает расти дочерний актиновый филамент. Таким образом и формируется древообразная структура, которая создает дендритные шипики, на основе которой и держится все волшебство нервной системы.
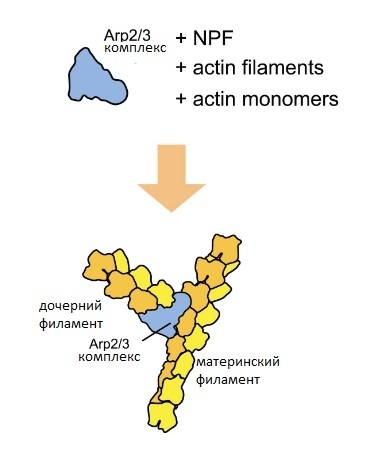
Изображение 2. Схема ветвления актиновых филаментов (адаптировано из (4)).
Тут уже не сложно догадаться, что если система дендритных шипиков нарушена, то можно ожидать нейро-когнитивныый дефицит, или же, по меньшей мере дисбаланс. На рисунке (изобр.[3]) показаны отростки нейронов дикого типа мыши (верхние изображения) и с прижизненно выключенным (нокаутированным) комплексом Arp2/3 (нижние изображения). Как можно заметить, существующие дендритные шипики у мутантов исчезают, а новые не появляются. Eсть исследования, которые показывают, что мутантнтные мыши, по комплексу Arp2/3 демонстрируют ненормальное локомоторное поведение (1). Более того, Arp2/3-мутантные мыши после введения антипсихотиков (галоперидол, клозапин) демонстрируют нормализацию поведения. А статья (2) обсуждает шизофрению в контексте изменения экспрессии гена Arp2/3.

Изображение 3. Динамика дендритных шипиков у мышей дикого типа (верхний ряд) и с инактивированным геном ArpC3 (нижний ряд) в течении восьми недель (адаптировано из (3)).
На рисунке (изобр.[4]) показаны данные, полученные с использованием анализа моторной активности мышей (общая длина передвижений) у мышей с интактным и инактивированным геном ArpC3 (это субъединица комплекса Arp2/3) с применением галоперидола (0.1 мг/кг, 0.2. мг/кг) и клозапина (0.5 мг/кг). Показаны три временные точки после введения антипсихотиков. В исследовании использовали Cre-LoxP систему инактивации гена, для простоты интерпретируем это как инактивированный ген (по-научному это генотип Arpc3 f/f; Camk2a-Cre).
Можно сразу заметить по первым двум столбцам, что у мышей с инактивированным геном ArpC3 общая дистанция передвижений значительно больше, чем у мышей с интактным геном (примерно в 6 раз). Анализируя диаграммы дальше можно заметить, что с течением времени антипсихотические средства сокращают разницу в моторной активности между мышами с интактным и инактивированным геном.
Ответ мутантных мышей на антипсихотические препараты объясняется тем, что потеря функциональной активности комплекса Arp2/3 непосредственно влияет на дофаминэргические синапсы сообщения вентральной области покрышки и черной субстанции (DA-producing VTA/SNc neurons).
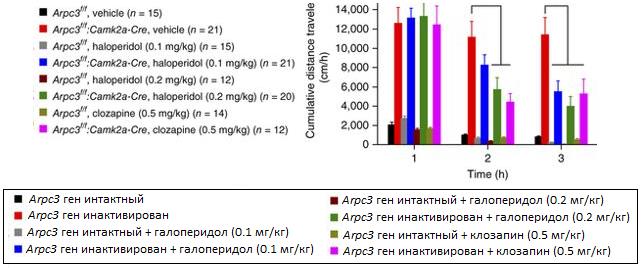
Изображение 4. Сравнение уровня моторной активности мышей дикого типа и с инактивированным геном ArpC3, без и с использованием антипсихотических средств, использованы три временные точки (адаптировано из (1))
На данный момент нет однозначных данных, описывающих биологическую сторону психической симптоматики, но регулярно появляется информация о новых генах, которые могут вносить вклад в ее развитие. Обзор о конкондартности шизофрении описывает наследуемость шизофрении на основании близнецовых исследований. В целом сомнений в генетической природе/компоненте психической патологии остается все меньше, однако пройдет еще много лет, прежде чем в психиатрическую практику войдут диагностические системы оценок рисков и предимплантационная диагностика. Расширение понимания биологических основ психиатрии существенно увеличит рецепторный арсенал психофармакологии, позволит проводить более селективную терапию и даже откроет возможности для доманифистационной коррекции в психиатрии.
Автор текста: Крат С.
Источники:
- Kim, Il Hwan, et al. “Spine pruning drives antipsychotic-sensitive locomotion via circuit control of striatal dopamine.” Nature neuroscience 18.6 (2015): 883.
- Datta, Dibyadeep, et al. “Altered expression of ARP2/3 complex signaling pathway genes in prefrontal layer 3 pyramidal cells in schizophrenia.” American Journal of Psychiatry 174.2 (2016): 163-171.
- Kim, Il Hwan, et al. “Disruption of Arp2/3 results in asymmetric structural plasticity of dendritic spines and progressive synaptic and behavioral abnormalities.” Journal of Neuroscience 33.14 (2013): 6081-6092.
- Helgeson, Luke A., and Brad J. Nolen. “Mechanism of synergistic activation of Arp2/3 complex by cortactin and N-WASP.” Elife 2 (2013): e00884.