
Для шизофрении и других психотических расстройств характерны галлюцинации, иллюзии и другие нарушения восприятия. Восприятие является субъективном процессом, посредством которого нейронные системы отображают и интерпретируют поступающие из внешней среды сенсорные сигналы. В силу того, что эти входящие сигналы всегда сопряжены с определенным уровнем «шума» или неоднозначности, способность сенсорных систем отсекать этот «шум» и вычленять конкретную необходимую информацию представляется ключевой функцией, обеспечивающей адекватную адаптацию к внешней среде.
Чтобы определить, что именно во входящем сенсорном потоке важно, сенсорные системы идут по наиболее оптимальному пути – обращаются к уже пережитому опыту. Тем не менее, даже эта «оптимальная» система обработки входящей информации не застрахована от ошибок. Как мы более подробно обсудим ниже, пережитый опыт подталкивает сенсорные системы к восприятию именно того, что уже было воспринимаемо раньше. А это, в свою очередь, приводит к ошибкам «восприятия-суждения» (perceptual-judgment errors), заставляя нас действовать по наиболее вероятному, но неверному в настоящий момент сценарию.
Такие ошибки довольно часто встречаются у здоровых людей и приводят к целой россыпи небольших обманов восприятия. Так, например, мы нередко можем видеть человеческие лица в облаках или чувствовать вибрацию от телефона в кармане, хотя нам никто не звонит.
Инкорпорирование прошлого опыта является уязвимой точкой процесса восприятия и, под воздействием определенных патологических процессов, может приводить к гораздо более выраженным нарушениям обработки сенсорной информации. В этом одна из причин такого широко известного психотического проявления, как слуховые вербальные галлюцинации, то есть восприятие пациентами голосов при отсутствии звучащей речи.
В основе предлагаемой нами концепции лежит идея о том, что в нарушениях восприятия, сопровождающих психоз, проявляется экстремальный вариант погрешностей в обработке сенсорной информации, которые естественном образом возникают в результате инкорпорирования предшествующего опыта.
Мы сделали выборочный обзор литературы, касающейся патофизиологии психотических нарушений восприятия, и попытались составить комплексную модель этих нарушений, тесно связанную с моделями нормального восприятия. По нашему мнению, эта концепция способна улучшить понимание этих загадочных феноменов. Мы постарались соединить воедино результаты исследований в нескольких областях, традиционно развивавшихся независимо друг от друга. Например, по результатам большого количества фармакологических и молекулярных исследований установлено, что центральную роль в развитии психоза играет избыточная дофаминовая передача в стриатуме. Однако предыдущие исследования обработки сенсорной информации при шизофрении концентрировались, в основном, на коре головного мозга и локальных путях в ней, практически игнорируя вклад нейромедиаторных нарушений и их влияние на функционирование нейронных сетей в базальных ганглиях.
Вероятно, это было связано с традиционным представлением о сенсорных системах как об изолированных отделах мозга, пассивно воспринимающих внешние стимулы и не зависящих от когнитивных влияний. В свою очередь, дофаминовые сети в стриатуме традиционно обсуждались в контексте системы награды и связанного с ней обучения, несмотря на накопленные доказательства важности стриатума и дофамина в когнитивных и перцептивных процессах в целом. Иными словами, роль стриатума истолковывалась слишком узко и ограничивалась процессами обучения, связанного с наградой, и процессом выбора действия. Лишь недавно появились данные, свидетельствующие о роли стриатума в процессах обучения сенсорных систем.
В нашей работе мы предприняли попытку объединить результаты последних исследований роли дофамина и базальных ганглиев в общую комплексную модель перцептивных нарушений, отталкиваясь от существующих теоретических моделей нормальных процессов восприятия. Учитывая широкую распространенность при психотических процессах нарушений именно слухового восприятия (по сравнению с нарушениями в других сенсорных модальностях), мы сконцентрировались на пересечении между дофамином, базальными ганглиями и системой обработки слуховой информации. Особое внимание уделено физиологии слухового восприятия и возможным патофизиологическим механизмам, которые могут привести к аудиторным нарушениям при психозе.
Феноменология
Пациенты, страдающие шизофренией или другими психотическими расстройствами, практически всегда сообщают о тех или иных нарушениях восприятия. Эти феномены, в частности, галлюцинации, на протяжении долгого времени тщательно фиксировались психиатрами и стали своеобразной «эмблемой» психической патологии в социокультурном пространстве. Как и другая продуктивная симптоматика, нарушения восприятия, как правило, начинают развиваться в молодом возрасте. В большинстве случаев они сначала, в продромальной препсихотической фазе, имеют неявные, аттенуированные формы. Затем, с прогрессированием психоза, они постепенно переходят в отчетливо выраженные галлюцинации.
Иллюзии и схожие феномены
Под иллюзиями понимают искаженное восприятие объективного реального стимула. Для психоза часто характерно иллюзорное восприятие действительности. Стимулы могут казаться измененными по своей интенсивности, формы объектов (например, лиц) также могут искажаться. Во многих случаях, особенно на ранних стадиях развития психоза, внешние стимулы приобретают в восприятии пациента более сильную, чем обычно, интенсивность. Соответственно, восприятие этих необычных стимулов приводит к необходимости их интерпретации и придания им какого-то значения. Этот период ощущения субъективной необычности собственных переживаний и поиска их смысла называется «бредовым настроением».
Галлюцинации
Галлюцинации – это ложное восприятие, никак не связанное с наличием объективного стимула во внешнем мире. Пациенты с шизофренией и расстройствами шизофренического спектра могут испытывать галлюцинации во всех сенсорных модальностях. Галлюцинации могут быть зрительными, слуховыми, обонятельными, соматосенсорными и даже вкусовыми. Однако именно слуховые галлюцинации наиболее распространены и наиболее выражены.
То, что, в отличие от других симптомов шизофрении, галлюцинации по своей природе дискретны (то есть могут «включаться» и «выключаться»), делает их крайне ценными в клиническом отношении. Хотя у некоторых пациентов, слуховые галлюцинации представляют из себя довольно простые звуки (например, звук шагов), у большинства преобладают галлюцинаторные голоса, проговаривающие не только отдельные звуки и слова, но и крайне сложные фразы, предложения, комментарии действий пациента, а также целые диалоги между несколькими знакомыми или незнакомыми голосами.
Содержание речи галлюцинаторных голосов может быть различным, но чаще всего это речь негативного содержания. Голоса могут оскорблять и унижать пациентов, часто ссылаясь на личный опыт пациента. При острых состояниях, вербальные галлюцинации, как правило, интерпретируются как реальные голоса, звучащие откуда-то извне. То есть пациент не осознает их как «внутренние» и «ненормальные» и действует соответствующе: громко им отвечает, пытается найти их источник и т. д.
Интересно, что при других формах психоза, например, при вызванном чрезмерным приемом продофаминергических препаратов в качестве терапии болезни Паркинсона, слуховые галлюцинации встречаются реже, чем зрительные. Этот факт дает возможность предположить, что система обработки именно слуховой информации в большей степени вовлечена в патофизиологию психотических нарушений при шизофрении и расстройствах шизофренического спектра.
Бред и психотический синдром
При психозе галлюцинации и другие перцептивные нарушения, как правило, развиваются параллельно с бредом. Под этим термином понимаются стойкие, не соответствующие действительности убеждения, сохраняющиеся даже при наличии прямых и явных доказательств их несостоятельности (и возникающие на болезненной основе – прим.ред.). Данное сочетание нарушений восприятия и бреда представляет собой особую клиническую конструкцию, определяемую как «психотический синдром».
Тем не менее, у небольшой части пациентов могут присутствовать только галлюцинации без бреда или наоборот. Исходя из этого, можно предположить, что наравне с общим патофизиологическим механизмом, обуславливающим развитие психотического синдрома как совокупности бреда и галлюцинации, существуют также и специфические пути развития каждого из симптомов по отдельности.
Облегчающие и отягчающие факторы
Данные огромного количества рандомизированных клинических исследований показали, что на выраженность психотического синдрома могут влиять препараты, блокирующие дофаминовые D2-рецепторы, наиболее плотно расположенные в стриатуме. Хотя у определенной когорты пациентов психотические симптомы могут продолжать развиваться и на фоне антипсихотической терапии, у большинства респондеров достигается клиническая ремиссия спустя несколько недель приема препаратов.
Важно отметить, что под клиническим улучшением подразумевается редукция продуктивной симптоматики, а не негативных симптомов или когнитивного дефицита. Это подчеркивает роль стимуляции D2-рецепторов в стриатуме в развитии психотической симптоматики, что впоследствии было подтверждено молекулярными исследованиями, обсуждаемыми ниже.
Имеются отдельные данные об уменьшении симптомов нарушения восприятия благодаря когнитивно-поведенческой терапии и транскраниальной магнитной стимуляции левой височно-теменной доли, но, в целом, никакие другие терапевтические подходы не показали своей эффективности в редукции психотической симптоматики. Стрессовые факторы внешней среды, наркотики (кетамин, каннабис и кокаин) и продофаминергические препараты (амфетамин) могут ухудшать и даже вызывать нарушения восприятия и другие психотические симптомы. Учитывая тот факт, что и стресс, и перечисленные выше препараты усиливают дофаминергическую передачу, а также данные об эффективности D2-антагонистов в лечении психоза, можно с уверенностью утверждать, что именно дофамин играет центральную роль в патофизиологии перцептивных нарушений и психотического синдрома как отдельной нейробиологической конструкции.
Такой вывод нисколько не умаляет роль других факторов (например, глутамат- или ГАМК-ергической передачи) в развитии психоза. Действительно, по некоторым данным, у 1/3 пациентов, особенно у тех, кто длительное время не получал терапию, антидофаминергические препараты не уменьшают интенсивность галлюцинаций. Тем не менее, ни один из множества разрабатываемых недофаминергических препаратов не показал эффективность в редукции нарушений восприятия при психозе.
Когнитивные механизмы
C точки зрения теории вычислительных процессов, модулирование восприятия и обработки входящей сенсорной информации на основе предшествующего опыта, иными словами, принятие перцептивного решения, можно представить как процесс, направленный на корректную идентификацию и классификацию входящих сенсорных сигналов, т. е. на снижение вероятности возникновения ошибок «восприятие-суждение». В этой части статьи мы сфокусируем внимание на описании гипотетических алгоритмов, посредством которых нейронные сети проводят эту операцию. При их описании мы основывались на большом объеме литературных данных о механизмах принятия решений в рамках системы восприятия. Также мы обсуждаем возможные нарушения этих алгоритмов, связанные с аномалиями восприятия. В следующей части, в соответствии с тремя уровнями анализа Марра, мы обсуждаем нейробиологическое значение данных процессов в нейронных сетях мозга, а также их нарушения при психозе.
Для понимания работы системы восприятия необходимо прояснить две концепции. Во-первых, для минимизации ошибок оптимальные перцептивные решения должны преодолеть неопределенность внешних стимулов (например, вычленить тихие звуки в шумной обстановке) и неопределенность внутренних репрезентаций этих стимулов (например, преодолеть проблему вариабельности сигнала от нейронов, кодирующих сенсорную информацию). Во-вторых, в реальных условиях стимулы практически никогда не действуют изолированно. Они всегда возникают на фоне других стимулов, которые могут предсказывать появление друг друга. Именно по этой причине при принятии перцептивных решений использование предшествующего опыта столь выгодно, ведь оно позволяет использовать контекстуальные связи между стимулами и, тем самым, снизить количество ошибок.
Теория распознавания сигнала
Представьте себе следующую ситуацию. Человек стоит на шумной станции метро и ждет поезд, чтобы, наконец, добраться до дома после тяжелого рабочего дня. Внезапно он слышит непонятный звук, как будто кто-то произнес его имя. Ему необходимо быстро решить, действительно ли кто-то произнес его имя или нет, то есть, является ли этот стимул настоящим сигналом или всего лишь шумом. В такой ситуации (подробнее см. Рис. 1), как утверждает теория распознавания сигнала, решение зависит от относительной вероятности того и другого исхода (сигнал имеется или, соответственно, отсутствует), причем вероятность оценивается на основе внутренней репрезентации этих стимулов. Это можно выразить в виде формулы, называемой логарифмическим отношением правдоподобия (log-likelihood ratio, LLR): LLR=log (P(доказательство|сигнал присутствует))/(P(доказательство│сигнал отсутствует))
Согласно этой модели, если сила доказательств за и против наличия в стимуле сигнала эквивалентна, то вероятность того и другого исхода одинакова, следовательно, LLR=0. Если же доказательств наличия сигнала больше, то LLR>0. Соответственно, если больше доказательств отсутствия в стимуле сигнала, то LLR<0.
Иными словами, если внешний стимул абсолютно двусмысленный и его восприятие (внутренняя репрезентация) крайне неоднозначно, то, находясь в условиях максимальной сенсорной неоднозначности, слушатель продемонстрирует некий «несмещенный» ответ. Этот ответ не будет смещаться в сторону восприятия стимула как сигнала, либо как шума. Такие смещения ответов по шкале доказательств образуют равновесие между «попаданиями» и «промахами» – верным и неверным определением сигнала с одной стороны, и корректным отклонением информационной значимости стимула и «ложными тревогами» с другой (см. Рис. 1)
Байесовский вывод
Как мы уже отметили выше, для преодоления неопределенности можно использовать контекстную информацию. Именно из контекста рождаются ожидания, включенные в процесс принятия решений в соответствии с Байесовскими принципами. Согласно им, предшествующее знание используется для оценки вероятности наличия в стимуле сигнала. Например, пассажир метро в нашем примере мог получить СМС от друга, в котором тот предупреждает, что тоже спускается на станцию. В таком случае увеличивается вероятность того, что друг действительно окликнет нашего пассажира по имени. Соответственно, процесс принятия решения будет смещен в сторону более вероятного события, т. е. увеличится вероятность того, что наш пассажир услышит в шуме свое имя (подробнее см. Рис. 1).
Информация для формирования предварительных ожиданий может поступать как из эксплицитной (осознаваемой нами, включающей в себя насыщенные чувственными характеристиками эпизоды из прошлого или абстрактную информацию), так и из имплицитной (неосознаваемой нами) памяти.
Байесовский вывод также можно считать своеобразным рецептом, позволяющим комбинировать два и более источника информации в условиях общей неопределенности. Данную схему часто называют «взвешивание достоверности». Если рассматривать систему восприятия, ключевыми источниками информации в ней являются предшествующие ожидания (источник 1) и вероятность сигнала в стимуле (источник 2). Эти два источника комбинируются между собой и образуют вместе новое, дополненное ожидание, которое уже непосредственно влияет на процесс принятия перцептивного решения.
Дополнение ожиданий по такому «рецепту» является, скорее, интуитивным процессом. В его основе лежит придание большего веса тому источнику информации, который представляется более надежным или менее неоднозначным. Из этого следует, что предшествующим ожиданиям придается больший вес в ситуациях максимальной сенсорной неопределенности (т. е. зашумления стимула), либо в тех ситуациях, когда предшествующие ожидания четко определены. Таким образом, в нашем примере пассажир с большей вероятностью услышит свое имя, если звуковой стимул был крайне неоднозначен (т. е. если вокруг него одновременно много разных шумов) и, в то же время, если пассажир был уверен, что на станции его встретит друг.
В реальном мире информация редко поступает вся целиком и сразу. Как правило, она все же приобретается постепенно, фрагмент за фрагментом. В примере со станцией пассажир не сразу примет решение, а, скорее, постарается получить больше информации из окружения, например, обернется и проверит, не приближается ли кто-кто к нему. В условиях такого постепенного поступления информации принцип Байесовского вывода обеспечивает постоянное обновление ожиданий, которое учитывается при оценке вероятности наличия сигнала в стимуле после поступления каждого нового фрагмента информации.
Этот процесс описывается моделями «аккумуляции связанных доказательств» (bounded evidence accumulation model). Одна из них, диффузионно-дрейфовая модель (drift diffusion model (DDM)), описывает процесс аккумуляции доказательств для принятия сенсорных решений. Этот процесс продолжается до тех пор, пока доказательства в пользу одного из возможных вариантов не накопятся в достаточной степени. В этой модели предшествующие ожидания выражаются в смещении стартовой точки и, соответственно, в необходимости меньшего количества доказательств для достижения одного из порогов. Помимо смещения стартовой точки предшествующие ожидания также частично влияют на скорость набора этих доказательств.
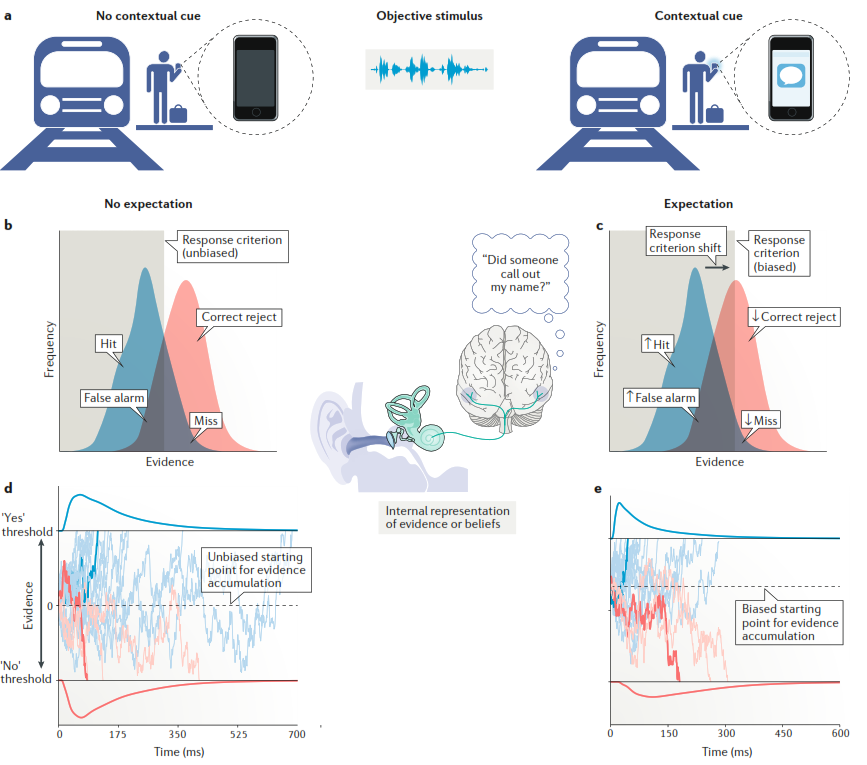
Рис. 1. Примеры принятия перцептивного решения при наличии или отсутствии «смещения ожидания», контекстно оправданной и адаптивной системы.
а. Эффект смещения, или предвзятости, ожидания можно объяснить на примере человека, ждущего поезд и слышащего неоднозначный звук (внешний объективный стимул (objective stimulus) в центре верхней части), напоминающий оклик по имени. В этом случае перцептивное решение (человек решает, услышал ли он действительно свое имя – решение «за» («yes» decision) – или не услышал – решение «против» («no» decision) – основывается на ожиданиях, отражающих внутренние представления о доказательствах за или против наличия в стимуле сигнала (оклик по имени).
b, c. Внешний объективный стимул начинает обрабатывается улиткой – органом, расположенным во внутреннем ухе. Улитка реагирует на звуковые вибрации и переводит их в нервные импульсы. В зависимости от силы сигнала увеличивается частота импульсации (на графиках показана по оси Y, «frequency»), идущей по слуховым путям в стволовые ядра, слуховые зоны таламуса и слуховую кору. В этих зонах – внутренние доказательства, ассоциированные с внешним стимулом (сила этих доказательств на графиках показана по оси Х, «evidence»). Синие гистограммы показывает силу доказательств, свидетельствующих за наличие сигнала в стимуле (оклик по имени). Красные гистограммы показывают силу доказательств против наличия сигнала. Серой зоной показан уровень, при котором происходит ответ на стимул (response criterion). Если человек перед этим неоднозначным звуком получает, например, СМС от друга на телефон, где тот сообщает, что тоже находится на этой станции (часть a, справа), то это смещает ожидание и, соответственно, критерии ответа на стимул, делая более вероятным распознавание в нем своего имени, то есть, сигнала, а не просто шума. Иными словами, увеличивается количество доказательств «за», что на графике c увеличивает зону, в которой произойдет ответ на сигнал. Это помогает корректно распознать сигнал (hit) и в то же время увеличивает вероятность ошибки, т. е. «ложной тревоги» (false alarm). Соответственно, в отсутствие этого предупредительного СМС (часть a, слева), нет смещения ожидания, зона ответа не изменяется (график b), и вероятность как верного ответа, так и «ложной тревоги» не изменяется.
d, e. Динамика накопления внутренних доказательств в соответствии с диффузионно-дрейфовой моделью (drift-diffusion model). Внутренние доказательства накапливаются до тех пор, пока доказательства за один вариант не наберут достаточной силы, чтобы преодолеть некий порог, после которого уже принимается соответствующее решение «за» или «против». Примерные траектории накопления доказательств «за» (синий цвет) и «против» (красный цвет) выделены на фоне других возможных траекторий (приглушенные цвета) при тех же модельных параметрах. Линии выше и ниже графика отражают распределение времени окончания процесса, ассоциированное с каждым из ответов (которое варьируется в силу достаточной «шумовой» насыщенности процессов накопления доказательств). На графике d (без предшествующего сигнала) начальная точка этого процесса накопления не смещена, на графике е, наоборот, смещена и расположена ближе к порогу ответа «за».
Предиктивное кодирование
Модели предиктивного кодирования математически сходны с моделями аккумуляции доказательств. Тем не менее, модели предиктивного кодирования описывают нейрональную архитектуру движения информации по разным уровням иерархического процесса обработки сенсорной информации. При этом более высокие уровни обработки способны создавать более сложные репрезентации.
В рамках моделей предиктивного кодирования аккумуляция доказательств представляется процессом обновления ожиданий, направленным на формирование моделей, объясняющих причины наблюдаемых сенсорных ощущений. Соответственно, при переносе информации по различным уровням системы обработки сенсорных сигналов уточняются распределения ожидаемых вероятностей наличия одной из двух возможных причин наблюдаемого сенсорного явления.
Основным алгоритмическим действием, повторяющимся на каждом иерархическом уровне, является интеграция нисходящих сообщений (т. е. ожиданий, сформированных из прошлого опыта) и восходящих сообщений, кодирующих, согласно данной модели, вероятность действительного наличия сигнала (ошибки ожиданий). Таким образом, модели предиктивного кодирования можно представить своеобразным сенсорным аналогом иерархических моделей подкрепления-обучения, в которых ключевым сигналом, предшествующим обучению, или же обновлению ожиданий, является ошибка ожидания не награды, а сенсорного переживания. Эти ошибки сенсорного ожидания взвешиваются согласно надежности получаемых восходящих и нисходящих сообщений. Соответственно, уже взвешенные ошибки сенсорного ожидания и определяют степень обновления ожиданий.
Суммируя все вышенаписанное, можно сказать следующее. Хотя разобранные концепции различаются, они обе строятся на предположении о том, что в процессе принятия сенсорного решения предшествующие ожидания встраиваются в новую сенсорную информацию посредством аккумуляции доказательств или обновления ожиданий.
Алгоритмические модели галлюцинаций
В 2005 г. исследователь Карл Фристон предложил алгоритмическую модель, в рамках которой галлюцинации являются чрезмерным смещением перцептивного процесса в сторону предшествующих ожиданий, возникающих из-за чрезмерной предшествующей уверенности, т. е. удельного веса ожиданий в процессе принятия перцептивного решения (см. Рис. 2). Он предположил, что галлюцинации – это перцептивный процесс, движимый в большей степени чрезмерным перцептивным ожиданием.
Основная предпосылка, на которой строится эта модель, аналогична Байесовским моделям лабораторных контекстно-обусловленных иллюзий. Однако модель Фристона предполагает, что смещение восприятия может происходить и при полном отсутствии объективного сенсорного стимула. Утверждение о том, что перцептивные решения определяют не только искаженное, но и ложное восприятие, может показаться несколько противоречащим здравому смыслу. Нужно иметь в виду, что изначальная вероятность наличия стимула в сигнале в Байесовской модели относится не к самому внешнему стимулу, а лишь к его внутренней репрезентации.
Так как сенсорные нейроны имеют определенный уровень спонтанной активности в отсутствие внешнего стимула, то получается, что в мозге никогда не существует полного отсутствия внутренних сенсорных доказательств. А это, в свою очередь, размывает границы между иллюзиями и галлюцинациями. Это очень интересный аспект этой модели. Галлюцинации в ней – это своего рода экстремальные иллюзии.
Этот вывод хорошо вписывается в представление о континууме нарушений восприятия и, в частности, объясняет постепенный переход искажений восприятия в полноценные галлюцинации при прогрессировании психоза. Особенно важно отметить то, что, согласно данной концепции, нейронные системы, ответственные за адаптивные перцептивные смещения, протекающие в норме, в патологических условиях способны обусловливать чрезмерные, дезадаптивные смещения, клинически выражающиеся как нарушения восприятия.
Более свежие дополнения к этой модели позволили определить ее в рамках иерархического предиктивного кодирования и возможности (аффорданса) действия. Согласно иерархическим моделям, существуют сложные взаимодействия между определенностью высокоуровневых предшествующих ожиданий и определенностью сенсорных доказательств на низших уровнях системы обработки сенсорной информации. Относительное увеличение определенности (иными словами, чрезмерная уверенность в высокоуровневых предшествующих ожиданиях по сравнению с низкоуровневыми сенсорными доказательствами) и вытекающие из этого перцептивные смещения и нарушения восприятия могут быть следствием компенсации диаметрально противоположных состояний.
Пациенты с шизофренией могут иметь изначальные более стабильные изменения, выражающиеся в относительном уменьшении уверенности в высокоуровневых предшествующих ожиданиях, что может компенсироваться уменьшением уверенности в низкоуровневых сенсорных доказательствах. Эта компенсация может сама по себе обусловливать галлюцинаторное психотическое состояние (см. Доп. 1 для информации об остальных теориях и моделях нарушений восприятия при психозе).
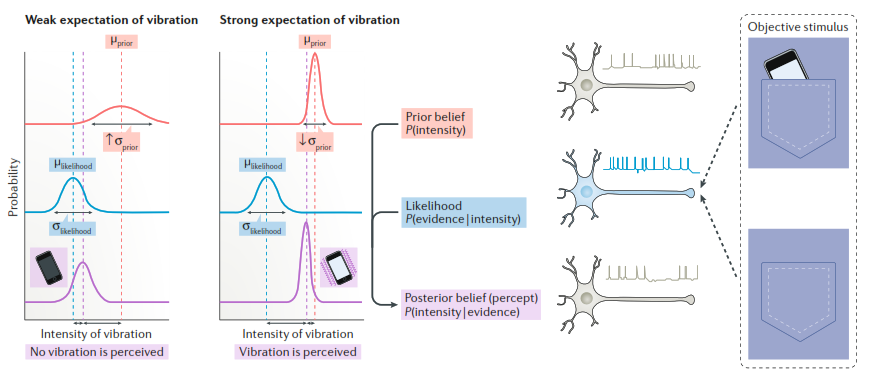
Рис. 2. Байесовская модель принятия перцептивного решения, иллюстрирующая смещение ожидания как при распространенных небольших искажениях восприятия, так и при выраженных галлюцинациях. Иллюстрация феномена «фантомной вибрации», который, несмотря на его широкую распространенность, можно рассматривать как галлюцинацию. Объективные внешние стимулы (либо легкое скольжение телефона в кармане по ноге (верхняя правая картинка), либо вообще отсутствие какого-либо вибротактильного стимула (нижняя правая картинка)) ассоциированы (стрелки) с внутренними сенсорными репрезентациями (доказательствами) в мозге – на схеме показаны как нейрон, кодирующий их в зависимости от частоты импульсации. Важно отметить, что частота этой импульсации, и соответственно, внутренние доказательства никогда не отсутствуют полностью: импульсация имеет место даже при полном отсутствии внешнего объективного стимула. При заданном уровне внутренних доказательств, конечное восприятие зависит от силы (однозначности или точности) предшествующих ожиданий. То есть, от того, насколько сильно мы ожидаем вибрацию телефона (например, ожидая звонок). Процесс обновления ожиданий, определяющий конечное, финальное восприятие, показан изогнутой стрелкой между выделенными разными цветами текстовыми блоками. Предшествующее ожидание (prior belief, выделено красным) комбинируется с вероятностью наличия сигнала (likelihood, выделено синим) и становится последующим, конечным ожиданием, и определяющим восприятие (posterior belief (percept), выделено фиолетовым). Слабое ожидание (левый график) характеризуется предшествующим ожиданием с высоким уровнем неоднозначности (высокое σprior); сильное ожидание (правый график) характеризуется низким уровнем неоднозначности (низкое σprior). Напротив, ожидаемая сила вибрации (μprior) ниже в первом сценарии и выше во втором. В обоих сценариях распределение изначальной вероятности уровня неоднозначности (σlikelihood) и ожидаемой силы вибрации (μlikelihood) в среднем одинаково. Финальное восприятие соответствует конечному ожиданию, которое в этих двух сценариях различно. При высоком уровне ожидания вибрация все же ощущается субъективно, т. к. конечное ожидание близко к предшествующему ожиданию и далеко от изначальной вероятности объективного наличия этой вибрации. При низком уровне ожидания, наоборот, конечное ожидание далеко от предшествующего ожидания и близко к изначальной вероятности, следовательно, вибрация субъективно не ощущается. Таким образом, при ожидании телефонного звонка фантомная вибрация телефона может появляться как при небольшом трении телефона о ногу в кармане (тогда это иллюзия), так и при полном отсутствии вибротактильной стимуляции (тогда это уже галлюцинация).
Поведенческие исследования галлюцинаций
Несмотря на то, что у пациентов, страдающих шизофренией, встречается довольно широкий набор изменений процессов восприятия, лишь некоторые из них, по-видимому, коррелируют с тяжестью галлюцинаций и других перцептивных нарушений. При рассмотрении галлюцинации в рамках модели детекции сигнала, можно сделать вывод, что они аналогичны «ложным тревогам». Было обнаружено, что при выполнении заданий, связанных с распознаванием звуковых сигналов в насыщенном шумами стимуле, пациенты, испытывающие слуховые галлюцинации, склонны слышать в этих стимулах речь и другие звуковые сигналы как при наличии, так и при отсутствии предварительных ассоциативных подсказок.
В нескольких исследованиях, где использовалась DDM (диффузионно-дрейфовая модель), были обнаружены нарушения алгоритмов аккумуляции доказательств при психозе. В частности, было показано, что болезнь Паркинсона со зрительными галлюцинациями ассоциирована с нарушением процессов сбора доказательств, которое выявляется в заданиях, связанных с визуальным восприятием. Имеются также предварительные данные, свидетельствующие о том, что при шизофрении несколько смещается стартовая точка процесса накопления доказательств. Это приводит к нарушению процесса интеграции предшествующих ожиданий, хотя и не совсем ясна связь этого параметра со зрительными галлюцинациями.
У лиц с повышенным риском развития психоза было обнаружено чрезмерное влияние предшествующих ожиданий на чувствительность восприятия при распознавании неоднозначных визуальных стимулов, но не на смещение ответа. Важно отметить, что этот эффект относится именно к начальным изменениям восприятия в субклинической популяции.
В другом исследовании было показано, что психотические пациенты, страдающие выраженными галлюцинациями, в задании по воспроизводству длины звукового тона чаще полагались на предшествующие ожидания, полученные из контекстных звуков, что определяло изменение субъективного восприятия целевого тона с неизменной длительностью. Наблюдаемый эффект был специфическим именно для галлюцинаций и сочетался с неспособностью изменить вес ожиданий в более вариабельных (неоднозначных) условиях. Поведенческие исследования, проведенные в парадигме Павловского обусловливания, показали, что пациенты, страдающие галлюцинациями, с различными клиническими диагнозами, в большей степени ожидают услышать тон после световой подсказки. Это также доказывает чрезмерный вес предшествующих ожиданий в принятии перцептивного решения у таких пациентов.
Таким образом, результаты представленных выше исследований согласуются с моделью галлюцинаций как результата чрезмерного влияния предшествующих ожиданий на восприятие. Некоторые из этих работ также в определенной степени доказывают основанное на клинических данных утверждение о наличии механизма, специфического для нарушений восприятия. Результаты этих исследований показывают, что обнаруженные особенности тесно связаны с галлюцинациями, а не с бредом или другими симптомами.
Также примечательно то, что обнаруженные корреляции между тяжестью галлюцинаций и наблюдаемыми во время лабораторных тестов признаками изменений восприятия подтверждают существование предложенного Фристоном континуума перцептивных нарушений. Возможным объяснением чрезмерного смещения восприятия при психотических перцептивных нарушениях может являться неспособность правильно встраивать данные о неопределенности окружения в предшествующие ожидания. Другой причиной может являться неправильная оценка определенности окружения, с приданием большего веса предшествующим ожиданиям даже в тех ситуациях, когда высокая неоднозначность контекста, окружающего стимул, делает эти предшествующие ожидания неинформативными. Но каков же все-таки биологический субстрат этих механизмов в головном мозге?
Нейробиологический субстрат
В следующей части работы мы сфокусируемся на перцептивных процессах, относящихся, главным образом, к слуховому восприятию. Однако сначала мы хотим обратить внимание на краткое описание литературных данных о процессе принятия решений при зрительном восприятии (см. доп. 2, рис. 3). Согласно этим данным, существует целая сеть, соединяющая кортикальные и субкортикальные регионы и ответственная за накопление доказательств в соответствии с диффузионно-дрейфовой моделью. Важно отметить, что интеграция предшествующих ожиданий выражается в смещении уровня базальной активности нейронов, «настроенных» на определенные особенности стимула и, соответственно, подающих сенсорные сигналы нижележащим ассоциативным зонам, ответственным за накопление доказательств.
Ниже мы приводим данные, свидетельствующие о том, что объединенные в единую сеть ассоциативные зоны коры, вовлеченные в обработку слуховой информации (ассоциативная слуховая кора и нижележащие ассоциативные зоны стриатума), играют важную роль в перцептивном смещении и развитии нарушений восприятия при психозе. Необходимо отметить, что, согласно современным моделям сознания, за осознанное восприятие ответственны скорее нижележащие ассоциативные зоны, а не сама сенсорная кора. Эти модели подразумевают наличие сопутствующей осознанному ответу на воспринимаемую информацию рекуррентной активности сети, включающей в себя ассоциативные зоны. Существование такой сети подтверждается исследованиями на приматах, в которых было обнаружено, что эти нижележащие ассоциативные зоны ответственны за осознанный перцептивный ответ (даже при «ложных тревогах»), а смещения их базальной активности ассоциированы со смещением ответа в ту или иную сторону.

Рис. 3. Нейробиологический субстрат механизма принятия решений в системе генерации саккад
Иллюстрация диффузионно-дрейфовой модели (DDM) процесса накопления доказательств при зрительном распознавании движения и ответственных за этой нейронных систем.
а. Стимулом в задании является облако случайно движущихся точек, целью же задания является точное определение преимущественного направления, по которому движутся точки. Эксперимент проводится следующим образом. Голова обезьяны фиксируется, затем на экране появляются две мишени: одна находится в рецептивном поле (receptive field, RF) нейрона, активность которого записывается, другая – в противоположной стороне. После этого появляется движущийся стимул, и обезьяна принимает решения, совершая саккадированные движения глазами в сторону одной из мишеней. Интенсивность наблюдаемого движения зависит от доли точек, движущихся в одном направлении (когерентности).
b. Анатомическое строение мозга приматов с выделенными зонами, ответственными за генерацию саккад. Средняя височная кора (MT, розовая) представляет из себя высокоорганизованную зрительную зону, чьи нейроны «настроены» на распознавание направления движения. Отростки от этих нейронов идут в нижележащие ассоциативные зоны (синие), ответственные за интеграцию доказательств (в том числе, в латеральную межтеменную кору (LIP) и хвостатое ядро, являющееся частью ассоциативных зон стриатума). В свою очередь, от этих зон идут отростки ниже, к зонам, напрямую управляющим глазными мышцами (зеленые) и, соответственно, вызывающим саккады. Стрелками указаны направления связей между зонами различных уровней.
с. Симуляция DDM-модели входящей функциональной единицы – модельного эквивалента нейронного ансамбля – генерирующей моментальное доказательство посредством изменения среднего ответа популяции нейронов на разную интенсивность сенсорных доказательств, ассоциированных со стимулом (слабая, средняя или сильная когерентность; график слева), а также ответа нижележащей функциональной единицы, аккумулирующей эти доказательства с течением времени (правая панель). Пунктирная линия обозначает время презентации стимула (stimulus on). Аккумуляция доказательств проявляется как резкое увеличение ответа от популяции нейронов, при этом чем сильнее доказательства, тем более резкий подъем.
d. Данные, полученные в ходе физиологических экспериментов на обезьянах при измерении ответов от единиц, соответствующих конкретным нейронным ансамблям. Было обнаружено, что активность зрительной зоны МТ (слева) соответствует моментальному доказательству, в то время как профиль активности сети, включающей LIP и хвостатое ядро (ассоциативный стриатум), соответствует аккумуляции доказательств во время презентации стимула (средний график). Для удобства показана только активность, соответствующая RF нейронов. Крайний правый график показывает, что наибольшие смещения базальной, престимульной активности как в LIP, так и в ассоциативном стриатуме (обозначенные звездочками в периоде, предшествующем презентации стимула), обладают наибольшим влиянием на процесс принятия решений (более высокую вероятность выбора направления, на которое «настроены» нейроны). Это особенно заметно при слабых стимулах, что соответствует Байесовскому принципу, говорящему о том, что смещение ожиданий должно наиболее сильно влиять в ситуациях наибольшей сенсорной неоднозначности.
Слуховая система
Отростки кохлеарных нейронов идут к ядрам ствола мозга, представляющим собой первый этап системы обработки слуховой информации (см. Рис. 1). Затем идут нейроны нижних холмиков, ядра медиальных коленчатых тел таламуса и первичная слуховая кора, у людей расположенная в поперечных височных извилинах (извилины Гешля). Эта первичная зона слуховой коры, как и на предшествующих уровнях, сохраняет тонотопическую организацию (принцип, согласно которому отдельные субпопуляции нейронов «настроены» на определенную частоту звука), отростки от нее отходят к близлежащим поясным и парапоясным зонам слуховой коры.
Вторичные и ассоциативные зоны слуховой коры располагаются вдоль верхней височной извилины. Паттерны ответов у этих зон сложнее, т. к. включают в себя распознавание человеческой речи и других видоспецифичных звуков, издаваемых голосом. Кроме того, эти зоны необходимы для распознавания эмоциональных (просодии) и неэмоциональных особенностей речи.
В классических исследованиях с участием пациентов, страдающих эпилепсией, было показано, что электрическая стимуляция ассоциативной слуховой коры, приводит к субъективному восприятию голосов (в том числе знакомых) и других сложных звуков у пациентов без психотической симптоматики. Кроме того, существует редкий подтип эпилепсии, затрагивающий ассоциативную слуховую кору таким образом, что во время приступа пациенты переживают слуховые галлюцинации, в том числе и вербальные.
У приматов ассоциативная слуховая кора, в отличие от первичной коры, имеет собственные эфферентные отростки, идущие к префронтальной коре и ассоциативным зонам стриатума (см. Рис. 4). К первичным и ассоциативным отделам слуховой коры подходят разные афферентные отростки. Ассоциативные отделы получают большее количество отростков от медиального коленчатого тела (кроме его первичного ретрансляционного ядра), а также от других ядер таламуса, включая медиальное ядро подушки.
Между уровнями обработки слуховой информации, в дополнение к перечисленным выше отросткам, обеспечивающим прямую связь, существует также густая сеть обратной связи, которая необходима для нисходящей регуляции перцептивных процессов в соответствии с иерархической архитектурой моделей предиктивного кодирования. Некоторые петли обратной связи включают в себя отростки, исходящие напрямую от латеральной префронтальной коры назад, к ассоциативной слуховой коре. Другие же могут идти по более извилистому маршруту через пути, соединяющие базальные ядра, таламус и кору.
Хорошо описаны схожие с ними пути в нижней височной коре приматов, ассоциированной с высокоуровневой обработкой зрительной информации и, возможно, играющей роль при развитии визуальных галлюцинаций. У грызунов также описаны схожие пути от базальных ганглиев к заднему ядру таламуса, а оттуда – в ассоциативные слуховые зоны коры, причем это те же зоны, что посылают отростки вперед, к ассоциативному стриатуму.
Все это дает возможность предположить, что замкнутая субкортикальная петля, ответственная за модуляцию высокоуровневых этапов обработки слуховой информации, действительно имеет место быть. Соответственно, эта петля является возможным нейробиологическим субстратом слуховых галлюцинаций (см. Рис. 4).
Изменения слуха при галлюцинациях
В фМРТ-исследованиях было показано, что для пациентов в психозе характерен определенный паттерн активности коры головного мозга, ассоциированный со слуховыми галлюцинациями. Если во время проведения исследования пациенты испытывали вербальные галлюцинации, то у них, как правило, выявлялась повышенная активность в ассоциативной слуховой коре, а также в верхней височной извилине и в зонах, расположенных поблизости к зоне Вернике. Некоторые исследования показали, что меняется даже базальная активность этих зон наблюдаемая в отсутствие галлюцинаций.
Все это свидетельствует о вовлеченности ассоциативной слуховой коры, ответственной за распознавание речи и других сложных звуковых конструкций, в развитие вербальных галлюцинаций при психозе. Более того, результаты подобных исследований приводят к интересному предположению о том, что усиление базальной активности этих воспринимающих речь зон может как раз-таки отражать смещение восприятия в сторону наиболее ожидаемого стимула – речи (см. Доп. 2).
Дофамин и сети базальных ганглиев
Одними из основных функций стриатума являются процессы принятия решений и выбора действия. Ключевым механизмом, посредством которого осуществляются эти процессы, является воротный механизм или, как его еще называют, гейтинг. Имеется в виду процесс активного контроля за переходом информации от одной структуры к другой. В базальных ганглиях гейтинг-механизм управляет таламо-кортикальным потоком информации, идущей от стриатума. Для поддержания этого механизма необходимы два пути: прямой (Go) путь контролируется средними игольчатыми нейронами стриатума (medium-spiny neurons, MSNs), экспрессирующими дофаминовые рецепторы 1-го типа (D1-MSNs). Их активация упрощает таламо-кортикальный поток информации и запускает поведенческую реакцию. Непрямой (NoGo) путь контролируется средними игольчатыми нейронами, экспрессирующими дофаминовые рецепторы 2-го типа (D2-MSNs), активация которых, наоборот, затрудняет таламо-кортикальный поток информации.
Дофаминовые нейроны среднего мозга – главным образом, в вентральной области покрышки (ventral tegmental area, VTA) и компактной части черной субстанции (substantia nigra, pars compacta, SNc) – имеют отростки, идущие в стриатум, сохраняя определенный топографический градиент; от VTA отростки идут по мезолимбическому пути в вентральный стриатум, а в различных зонах SNc – по нигростриарному пути в дорсальный стриатум (включая его ассоциативные и сенсомоторные зоны). В стриатум также идут отростки от корковых структур, сохраняя топографическую организацию. При этом они образуют синапсы с MSNs, создавая, таким образом, частично разделенные кортико-стриато-таламо-кортикальные петли. В ассоциативный стриатум идут отростки от различных ассоциативных корковых зон, причем эти входящие влияния частично перекрещиваются, образуя небольшие зоны конвергенции. Особенно много таких зон в переднем хвостатом ядре.
Фазовое высвобождение дофамина в стриатуме играет центральную роль в подкреплении обучения путем постепенного налаживания этой сети на основе переживаемого опыта. На клеточном уровне быстрые колебания уровня дофамина (dopamine transients) модулируют пластичность кортико-стриарных синапсов, включая долговременную потенциацию D1-MSNs и долговременную депрессию D2-MSNs. В дополнении к постепенным изменениям пластичности, изменение уровня дофамина модулирует возбудимость этой сети прямо во время принятия решений. Дофамин усиливает возбудимость D1-MSNs и снижает возбудимость D2-MSNs, эффективно регулируя приток входящей информации от стриатума.
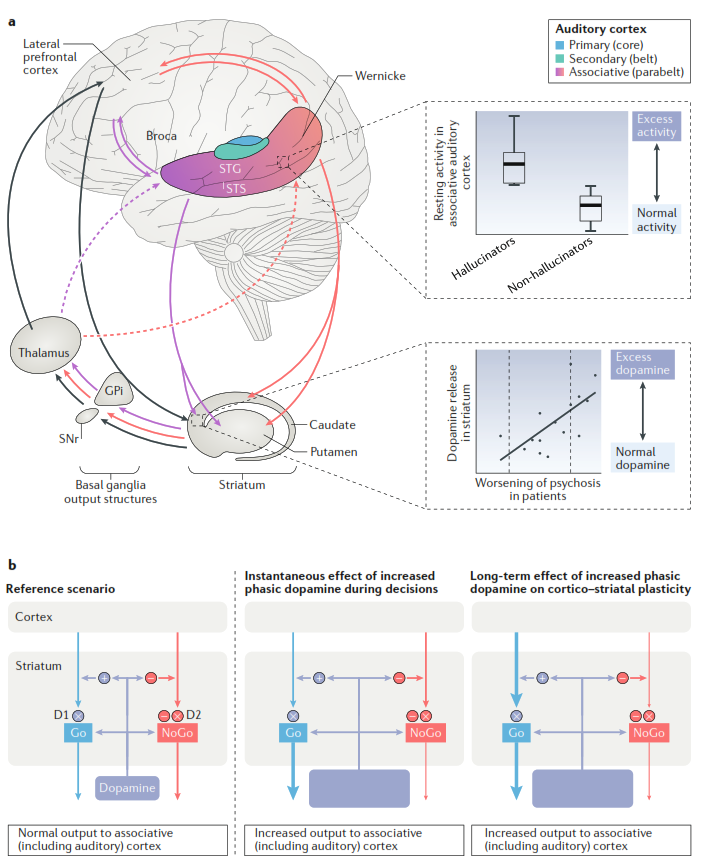
Рис. 4. Сети ассоциативной слуховой коры и базальных ганглиев, вовлеченные в формирования нарушений восприятия при психозе
а. Анатомия слуховой коры и нисходящих отростков от ассоциативной слуховой коры до префронтальной коры и стриатума, а также кортико-кортикальных и кортико-базально-таламо-кортикальных петель, предположительно играющих роль в развитии перцептивных нарушений. В центральной части показана латеральная поверхность мозга, на которой выделены разными цветами отделы слуховой коры. Функции нейронов в разных отделах ассоциативной слуховой коры по своим функциям неодинаковы. Таким образом, образуется своего рода градиент от передне-вентральной до задне-дорсальной части ассоциативной слуховой коры (выделено цветом). Отростки от слуховой ассоциативной коры идут в дорсальный стриатум и латеральную префронтальную кору (на рисунке показаны стрелками, цвет которых совпадает с цветом зоны слуховой ассоциативной коры, откуда они идут). Стрелки черного цвета также соответствуют входящим влияниям в префронтальную кору от других зон головного мозга. Для большей наглядности стриатум, таламус и базальные ганглии вынесены на рисунке отдельно. Также показаны связи, идущие от стриатума к базальным ганглиям и таламусу, а также таламо-кортикальные обратные связи. Пунктиром выделены связи, не описанные в достаточной мере у приматов. Отдельно показаны графики, иллюстрирующие аномальную активность ассоциативной слуховой коры при галлюцинациях и увеличение уровня дофамина при психозе.
b. Схематические изображения гипотетических изменений в кортико-базально-таламо-кортикальных сетях при шизофрении вследствие избытка дофамина. Слева показан нормальный, референсный, сценарий – нормальные уровни дофамина (темно-синий прямоугольник) и нормальный баланс путей средних игольчатых нейронов стриатума D1-MSN (Go, голубой) и D2-MSN (NoGo, красный). На средней и правой панелях показаны гипотетические варианты смещения баланса в сторону пути «Go» вследствие быстрых колебаний уровня дофамина (большие синие прямоугольники) при психозе. На средней панели показан кратковременный, мгновенный эффект быстрого увеличения уровня дофамина, активирующего «Go»-путь и ингибирующего «NoGo»-путь, воздействуя напрямую на стриарные MSNs (обратите внимание на неизмененные кортико-стриарные входящие влияния). На правой панели показаны долговременные эффекты повышенного уровня дофамина на кортико-стриарную пластичность, приводящие к усилению влияний на «Go»-путь и, наоборот, угнетению «NoGo»-пути. Оба аномальных сценария (на средней и правой панели) в конечном итоге приводят к усилению передачи от базальных ганглиев через таламус к ассоциативной слуховой коре.
GPi – internal globus pallidus (внутренний бледный шар); SNr – substantia nigra, pars reticulata (сетчатая часть черной субстанции); STG – superior temporal gyrus (верхняя височная извилина); STS – superior temporal sulcus (верхняя височная борозда).
Важная роль дофаминового сигналинга в сенсорных процессах предполагалась давно, однако только в последнее время появились достоверные данные, свидетельствующие о том, что дофамин участвует не только в процессах обучения, ассоциированного с наградой, но и в процессах сенсорного обучения. Известно, что неоднозначные зрительные и слуховые стимулы вызывают всплеск активности дофаминовых нейронов. Было также показано, что ответ этих нейронов отражает перцептивную неоднозначность таким образом, что его невозможно объяснить только ожиданиями награды.
Более того, в исследованиях на нечеловекообразных обезьянах было показано, что субъективное восприятие в отсутствие сенсорных стимулов ассоциировано с дофаминовым сигналингом. В другом исследовании, проведенном на грызунах, было показано, что быстрые изменения уровня дофамина являются необходимым и достаточным условием для возникновения ассоциаций стимул-стимул, даже если эти ассоциации формируются между нейтральными стимулами, не связанными с механизмами награды. В другом исследовании на грызунах с прямой оценкой быстрых колебаний уровня дофамина было показано, что, хотя дофаминовые сигналы в вентральном стриатуме соответствуют ошибкам ожидания награды, дофаминовые сигналы в некоторых отделах дорсального стриатума, наоборот, соответствовали ошибкам сенсорного ожидания, кодирующим непредсказуемость стимула.
Взятые все вместе, данные этих исследований свидетельствуют о том, что, помимо своей известной роли в сигналинге, связанном с ошибкой предсказания награды, быстрые колебания уровня дофамина могут кодировать не связанную с наградой информацию, относящуюся к перцептивным процессам.
Исследования с использованием про-дофаминергических препаратов у людей также свидетельствуют о роли дофамина в принятии перцептивных решений. В одном исследовании, посвященном болезни Паркинсона, было показано, что для пациентов, принимающих леводопу, в отличие от тех, кто ее не принимает, характерно снижение аккумуляции доказательств (сниженная скорость дрейфа в модели DDM) во время принятия перцептивных решений при выполнении задания со случайным движением точек (см. Доп. 3). Это свидетельствует о том, что относительное увеличение уровня дофамина приводит к меньшему влиянию сенсорных доказательств на принятие перцептивного решения.
В другой работе с использованием теста «соответствие силе» (force-matching) было показано, что с увеличением дозы леводопы ассоциировано большее влияние собственных сенсомоторных ожиданий и, соответственно, меньшее влияние сенсорных доказательств, что согласуется с результатами указанной выше работы. Тем не менее, в другом исследовании прием леводопы пациентами с болезнью Паркинсона, наоборот, приводил к увеличению «веса» сенсорных доказательств относительно предшествующих ожиданий в задании, связанном со зрительно-пространственными суждениями.
Еще одно исследование, проведенное при участии здоровых испытуемых, показало, что увеличение дофамина в стриатуме коррелировало с уменьшением сигналов обновления ожиданий, относящихся к выполнению задания. Таким образом, эти результаты согласуются с ролью дофамина при принятии перцептивных суждений, а также дают возможность предположить, что дофамин по-разному влияет на разные кортикостриарные системы.
Дофаминергические изменения при психозе
Дисфункция дофаминовых систем играет центральную роль в появлении и развитии нарушений восприятия и других психотических симптомов при шизофрении. Молекулярно-визуализационные исследования in vivo позволили изучить изменения плотности дофаминергических (особенно 1-го и 2-го типов) рецепторов, дофаминовых переносчиков и, опосредованно, уровня дофамина. В этих исследованиях фармакологически индуцировали резкие изменения уровней синаптического и перисинаптического дофамина, и затем изучали влияния этих изменений на захват D2-радиометок. Эти изменения являлись прямым следствием конкуренции между дофамином и радиометкой за связывание с рецептором, но также могли быть следствием перемещения (трафика) D2-рецепторов в нейронах после воздействия агониста.
Молекулярные исследования показали, что плотность D1-рецепторов и дофаминовых транспортеров в стриатуме не меняется. Плотность же D2-рецепторов в стриатуме была выше только у тех пациентов, кто получал антипсихотическую терапию, но не у пациентов, не принимавших препараты. Кроме того, было показано, что при психозе имеет место увеличение выброса дофамина в стриатуме, повышение уровня интрасинаптического дофамина, а также увеличение скорости его синтеза. Эти изменения особенно отчетливо заметны в ассоциативном стриатуме.
Более того, избыток дофамина в ассоциативном стриатуме связан с тяжестью психоза, в том числе с тяжестью галлюцинаций, как у пациентов с установленным диагнозом, так и у испытуемых из группы высокого риска развития заболевания. Даже у пациентов, страдающих шизофренией с коморбидной зависимостью, у которых обычно снижается общий уровень дофамина в стриатуме, все еще имеется эта корреляция между степенью повышения уровня дофамина и тяжестью психоза. Избыток дофамина в стриатуме также можно рассматривать как ранний предиктор развития психотических состояний. Избыточный дофамин характернее для острых состояний, нежели чем для более стабильных. Кроме того, он может возникать у пациентов с аффективными психозами, не страдающими шизофренией.
Можно предположить, что разбираемый фенотип характерен в большей степени для психотического синдрома, нежели чем для шизофрении как цельной нозологии. Изменения дофаминовой передачи в стриатуме, таким образом, являются наиболее достоверным нейробиологическим коррелятом психоза и, в частности, характерных для него нарушений восприятия.
Стоит также отметить, что в более ранних молекулярных исследованиях было показано, что существует определенная дозозависимая связь между процентом «занятых» D2-рецепторов и клинической эффективностью антипсихотиков, что, в сочетании с результатами множества клинических исследований, позволяет с уверенностью говорить о несомненной роли D2-рецепторов в терапии психотических состояний.
Собранные данные, во-первых, поддерживают идею о вкладе избыточного пресинаптического дофамина в развитие психоза, а во-вторых, дают возможность предположить дополнительную роль постсинаптических D2-путей стриатума. Тем не менее, несмотря на явную вовлеченность стриарного дофамина в развитие психоза, точная природа дофаминовых нарушений требует дальнейшего изучения.
Дисфункция стриатума при психозе
Известно, что острые нарушения мозгового кровоснабжения (инсульты), затрагивающие стриатум, клинически могут проявляться перцептивными нарушениями, сходными с таковыми при психозах. В литературе есть сообщения о нескольких случаях галлюцинаций у пациентов с поражением дорсального стриатума. Кроме того, в некоторых фМРТ-исследованиях показано, что галлюциноз ассоциирован с увеличением метаболической активности дорсального стриатума, хотя в визуализационных исследованиях, оценивающих изменения кровотока на более коротком промежутке времени, подобных особенностей обнаружено не было.
Дофаминовая модель психоза
Согласно последней модели дофаминовых нарушений при шизофрении, изучение фазовых изменений дофамина (резкие увеличения его уровня), возможно, в сочетании с тоническим (постоянным) повышением уровня дофамина, поможет объяснить наблюдаемые в визуализационных и поведенческих исследованиях феномены через призму теории подкрепления-обучения. В основу этой модели положена двойственность эффекта дофаминовой дисрегуляции: с одной стороны, происходят долговременные изменения подкрепления-обучения вследствие изменений кортико-стриарной пластичности; с другой стороны – кратковременные изменения во время принятия решений посредством мгновенной модуляции возбудимости MSN (как это обсуждалось выше).
Высказывалось предположение, что дофаминовая дисрегуляция в стриатуме смещает баланс между D1- и D2-путями, долговременно потенцируя первый и ингибируя второй путь. Основным эффектом этого смещения является облегчение передачи от базальных ганглиев к корковым структурам («Go» механизм, гейтинг). Учитывая тот факт, что сети базальных ганглиев ответственны за гейтинг моторной, сенсорной и когнитивной информации посредством кортико-базально-таламо-кортикальных петель, данная модель предполагает, что именно облегчение прохождения входящих влияний из стриатума в слуховую кору и является потенциальным механизмом развития слуховых галлюцинаций.
Взаимодействие слуховой коры и базальных ганглиев
У приматов от ассоциативной слуховой коры отходят моносинаптические связи к стриатуму, в том числе к ассоциативным регионам в дорсальной его части. Это свидетельствует о вовлеченности ассоциативного стриатума в высокоуровневые процессы принятия перцептивного решения, о чем свидетельствуют теоретические изыскания и последние нейрофизиологические исследования на нечеловекообразных приматах и грызунах.
Обнаружено, что постоянная активность в ассоциативном стриатуме связана с процессом накопления доказательств при принятии перцептивного решения в заданиях, основанных на зрительном и на слуховом восприятии. Базальная (престимульная) активность нейронов, «настроенных» на то или иное перцептивное решение, определяет, какая из двух альтернатив будет выбрана при последующих перцептивных решениях. Особенно сильно это заметно при слабых сенсорных доказательствах, что согласуется с реализацией оптимального включения предшествующих ожиданий в перцептивные решения согласно Байесовским моделям (см. Рис. 3d)
У грызунов ассоциативный стриатум в контексте принятия слуховых перцептивных решений изучен хуже. Тем не менее, в последние годы все больше внимания уделяется так называемому «слуховому стриатуму» – дорсальной части его сенсомоторной зоны, получающей отростки от первичной слуховой коры и дофаминовых нейронов среднего мозга. Нейроны «слухового стриатума» участвуют в принятии слуховых решений и достаточно стабильно кодируют информацию о конкретных чертах слухового стимула (например, о частоте звука) даже в отсутствие сигналов, кодирующих реакции ожидания награды. Более того, было показано, что обучение в контексте слуховых решений зависит от пластичности слуховых кортико-стриарных связей, идущих к этой зоне, особенно нейронов, «настроенных» на относящиеся к заданию особенности звукового стимула (например, на высокую частоту, предсказывающую получение награды).
У людей ассоциативный стриатум также вовлечен в процессы принятия решений, что следует из результатов фМРТ-исследований. Активация стриатума во время выполнения задания со случайным движением точек коррелировала с появлением правильных прогнозирующих сигналов, следовательно, эта структура может играть роль во встраивании предшествующей информации, определяющей ответ на стимул. В одном исследовании было показано, что сигналы стриатума соответствуют уровню неопределенности в предшествующих ожиданиях при выполнении задач по зрительно-пространственному распознаванию.
Аудиторно-стриарные изменения при психозе
Существуют предварительные данные клинических фМРТ-исследований, предполагающие связь нарушений восприятия при психозе и дисфункции стриатума. В некоторых исследованиях было показано, что сети, включающие в себя стриатум, вовлечены в процессы принятия перцептивных решений у галлюцинирующих пациентов. Причем у этих пациентов было снижено количество предикторных сенсорных сигналов в скорлупе, что коррелировало со снижением предупреждающих об ошибке сигналов при выполнении задания на распознание речи.
В другом исследовании с применением парадигмы «условно-рефлекторных галлюцинаций» (conditioned-hallucination paradigm) было показано, что у галлюцинирующих пациентов смещался поведенческий ответ в сторону большей частоты «ложных тревог», при этом во время таких ответов регистрировалась активация стриатума. Повышение уровня дофамина в ассоциативном стриатуме коррелировало с тяжестью галлюцинаций, а также с невозможностью уменьшить влияние предшествующих ожиданий на принятие решения в задаче по воспроизводству длительности тона у пациентов с шизофренией, не получавших терапию.
Потенциальная комплексная модель
Учитывая все приведенные выше находки и опираясь на предыдущие модели, мы предполагаем, что возможным нейробиологическим механизмом развития нарушений восприятия при психозе являются нарушения процессов принятия перцептивных нарушений, связанные с ассоциативным стриатумом и уровнями дофамина. Мы предполагаем, что это достаточный, но не необходимый механизм, задействованный, по крайней мере, в некоторой части наблюдаемых при шизофрении галлюцинаций. Этот механизм вряд ли сможет объяснить все связанные с галлюцинациями феномены, наблюдаемые в клинических и в субклинических популяциях. Тем не менее, важно отметить, что описанный дофамин-зависимый механизм биологически оправдан, понятен в алгоритмических терминах и учитывает данные о роли дофаминовых нарушений стриатума в развитии психоза.
Как нами было обозрено выше, связанные с принятием перцептивного решения модели галлюцинаций предполагают чрезмерное влияние предшествующих ожиданий на перцептивные процессы, приводящее к восприятию ожидаемого и уводящее от восприятия реального и объективного. В случае с вербальными галлюцинациями, их появление можно объяснить усилением базальной активности специфических для восприятия речи нейронов ассоциативной слуховой коры, нисходящие отростки которых идут в ассоциативный стриатум и префронтальную кору (схожие данные, но для другой модальности восприятия, обсуждаются в Доп. 2). Это усиление может смещать процесс обновления ожиданий, идущий в ассоциативном стриатуме, в сторону распознавания в стимуле речи, что, в конечном итоге, приводит к росту числа «ложных тревог», когда стимул распознается как речь даже при ее отсутствии (а иногда и при отсутствии слухового стимула как такового, что и будет вербальным галлюцинозом; см. Рис. 4).
Хотя эта модель и предлагает объяснение лежащих в основе вербальных галлюцинаций смещений ожидания, она пока что не объясняет, чем же вызвано это усиление базальной активности селективных «речевых» нейронов слуховой коры. Однако эта модель может, например, помочь с пониманием того, как слуховая гиперреактивность возникает вследствие избытка дофамина в ассоциативном стриатуме, что было нами постулировано.
На ранних стадиях развития психоза быстрые увеличения уровня дофамина, отражающие высокий уровень перцептивной неоднозначности или ошибки сенсорного ожидания, могут приводить к состоянию субъективно воспринимаемой неоднозначности, «непонятности» входящей сенсорной информации, что клинически выражается в так называемом бредовом настроении. Гипердофаминергические состояния приводят к ошибочному приписыванию значимости нейтральным стимулам, усиливая готовность связывать между собой ничего не означающие события. У пациентов с развивающимися вербальными галлюцинациями эти усиленные дофаминовые влияния могут случайным образом усилить кортико-стриарную пластичность и вызвать дисбаланс между D1- и D2-путями. Этот дисбаланс приводит к долговременной потенциации D1-MSN-путей стриатума, по которым идут селективные для восприятия речи влияния, и, наоборот, к долговременному угнетению D2-MSN-путей. Подробные механизмы этого явления были описаны выше.
Происходящее – не результат случайности, а следствие того, что усиленная дофаминергическая передача в первую очередь вовлекает ту часть ассоциативного стриатума, которая собирает на себе отростки от селективно воспринимающих речь нейронов ассоциативной слуховой коры. С другой стороны, стрессовое состояние, ассоциированное с бредовым настроением, может само по себе предрасполагать пациента к поиску персонально значимой, часто негативной, интерпретации нейтральных стимулов. Это приводит к усилению активности нейронов, «настроенных» на негативно окрашенную (например, уничижительную) речь. Подобное усиление может происходить посредством нисходящих префронтальных влияний, «отбирающих» информацию только от этих нейронов, «настройка» которых соответствует предшествующим ожиданиям. Несмотря на это, увеличение объема дофаминовых сигналов может усилить входящие «речевые» влияния в ассоциативный стриатум и способствовать усилению передачи от базальных ядер к ассоциативной слуховой коре посредством дисбаланса между D1- и D2-путями. А это, в свою очередь, приводит к усилению базальной активности нейронов в распознающей речь области ассоциативной слуховой коры.
Это предположение подтверждается результатами предварительных исследований на грызунах, а также, в некоторой степени, визуализационными исследованиями нарушений дофаминовой передачи в стриатуме при психозе. Таким образом, усиливая базальную активность распознающих речь нейронов, а также нарушая процессы обновления ожиданий в стриатуме, селективная долговременная потенциация входящих «речевых» влияний приводит к длительному смещению восприятия в сторону распознавания в слуховом стимуле речи и, соответственно, к хроническому вербальному галлюцинозу.
Кроме того, в течение конкретного перцептивного события дофаминовые сигналы могут сообщать о перцептивной неопределенности путем модулирования скорости (путем мгновенных изменений относительной возбудимости D1-MSN по сравнению с таковой у D2-MSN) связанных с речью кортико-стриарных входящих влияний относительно других влияний. Говоря алгоритмическим языком, это и есть процесс взвешивания предшествующих ожиданий. При психозе избыточность и нечеткость дофаминовых сигналов может в зависимости от контекста уменьшать динамический диапазон кодирования перцептивной неопределенности. Это приводит к тому, что пациенты будут чересчур сильно полагаться на предшествующие ожидания даже в таких неоднозначных условиях, в которых само использование предшествующего опыта крайне неинформативно.
Эта модель, конечно, спекулятивна, однако у нее есть ряд привлекательных моментов. Во-первых, она одновременно объясняет и само смещение ожиданий, и неспособность его изменения даже при условии крайне неоднозначного контекста. Во-вторых, хотя согласно этой модели оба данных процесса и определяются дофаминовыми нарушениями в стриатуме, само смещение ожидания может возникать постепенно, как следствие изменяющихся кортико-стриарных связей, что, вероятно, соответствует постепенному переходу от иллюзий к галлюцинациям с развитием психоза. В то же время недостаточность контекстно-зависимых изменений этого смещения может быть более тесно связана с собственно уровнями дофамина.
Появляется возможность объяснить наше первоначальное наблюдение о том, что только недостаточность регулирования смещения ожиданий, а не сама величина этого смещения, коррелируют с увеличенными уровнями дофамина в ассоциативном стриатуме у галлюцинирующих пациентов. Также появляется возможность объяснить бимодальную временную кривую ответа на антипсихотическую терапию, которая, блокируя D2-рецепторы и, соответственно, усиливая кратковременную активность и долговременную пластичность D2-MSN путей, выравнивает тем самым баланс между D1- и D2-путями в стриатуме. Это обеспечивает кратковременную нормализацию предвзятости ожиданий, что может уменьшить выраженность галлюцинаций до определенной степени. Помимо этого, антипсихотики вызывают долговременные изменения кортико-стриарной пластичности, которые способны в полной мере нормализовать предвзятость ожиданий и, соответственно, убрать галлюцинации.
Кроме того, эта модель позволяет объяснить, почему антипсихотическая терапия работает менее эффективно в отношении некоторых пациентов с длительно протекавшим психозом, но не получавших лечение. У таких пациентов, предположительно, ассоциированные с психозом нарушения кортико-стриарной пластичности развились в такой степени и укоренились настолько глубоко, что скорректировать их медикаментозно представляется крайне трудной задачей, особенно если психотические нарушения возникли в раннем возрасте. Представленная патофизиологическая модель не предлагает прямого объяснения этиологии дофаминовой дисрегуляции, но можно предположить, что за развитие дисрегуляции отвечает комбинация генетических и средовых факторов, в том числе и связанных со стрессом, затрагивающих высокоуровневые структуры, регулирующие функцию стриатума и среднего мозга.
Модель можно расширить и попытаться объяснить бред как следствие изменений, с алгоритмической точки зрения, сходных и тесно связанных с таковыми при галлюцинациях. Вероятно, эти изменения приводят к формированию чрезмерной, ригидной уверенности в убеждениях, затрагивающих некие абстрактные, неявные феномены. За возникновение бредовых убеждений, скорее всего, ответственны соответствующие нейронные сети, которые в некоторой степени связаны с сетями, ответственными за возникновение галлюцинаций. С одной стороны, это объясняет частое сочетание этих симптомов, а с другой стороны, это объясняет различия между ними и возможность присутствия одного симптома в отрыве от другого. Предварительные данные последних исследований свидетельствуют о правильности этого вывода.
Тем не менее, остаются открытыми вопросы о том, как согласовать эту модель с огромным объемом данных о предполагаемой роли ошибок по типу поспешных выводов (jumping-to-conclusions), имеющих место при шизофрении, и о том, какие конкретно нейробиологические структуры представлены в различных звеньях построенной модели. Безусловно, потребуется большой объем работы для того, чтобы сравнить нашу модель, сфокусированную на нигростриарных дофаминовых нарушениях процессов принятия перцептивного решения, с другими моделями, предполагающими роль дофаминовых нарушений либо в системе награды, либо в системе предиктивных моторных сигналов, используемых для пометки действий как своих собственных.
В конце концов, как мы заявили ранее, мы предполагаем, что дофаминовая дисфункция стриатума является лишь одним из возможных механизмов развития нарушения восприятия. Сходные фенотипы могут проявляться и вследствие других, недофаминергических механизмов (см. Доп. 3).
Заключительные ремарки
В данной работе мы посмотрели на психотические нарушения восприятия с точки зрения алгоритмов принятия перцептивного решения и описали возможный дофаминергический механизм в стриатуме, обуславливающий предвзятость восприятия и имеющий отношение к перцептивным нарушениям. Недавние исследования свидетельствуют о ключевой роли дофамина в процессах перцептивного обучения и принятия решений, не связанных с системой награды. Исследования с приматами и грызунами подчеркнули роль ассоциативного стриатума в принятии перцептивных решений и в смещении восприятия в сторону предшествующих ожиданий.
Учитывая важность предвзятости ожиданий в современных моделях восприятия, а также данные о дофаминергических изменениях в ассоциативном стриатуме при психозе, в своей модели психотических нарушений восприятия мы ставим во главу угла смещение ожиданий как основной алгоритмический процесс, рассматривая переизбыток дофамина в стриатуме как основной нейробиологический субстрат этого процесса. Хотя нам остается ответить еще на множество важных вопросов о патофизиологии перцептивных нарушений при психозе, мы утверждаем, что именно такие теоретически обоснованные, комплексные подходы позволят добиться понимания такого загадочного феномена как галлюцинации, существование которых будоражило умы бесчисленных поколений людей.
Мы верим, что разработка и постоянная доработка теоретически обоснованных моделей, соединяющих биологию и субъективный опыт пациентов, является ключом к пониманию психоза и, безусловно, помогает дестигматизировать это состояние и разработать новое поколение лекарственной терапии.
Доп. 1. Модели нарушений восприятия при психозе
Теории «снизу-вверх». Высказывалось предположение, что за развитие перцептивных нарушений ответственны искаженные сенсорные сигналы, возникающие на ранних этапах обработки сенсорной информации. Это утверждение строится на предположении о том, что «чистые» входящие сенсорные сигналы, не измененные нисходящим модулированием, являются анатомически разобщенными на ранних стадиях обработки сенсорной информации, в отличие от множества нисходящих модуляторных воздействий и обратных связей на этих этапах. Соединяющим звеном теорий «снизу вверх» и «сверху вниз» может быть тот факт, что чрезмерная сенсорная неоднозначность может вторично приводить к избыточному увеличению «веса» предшествующих ожиданий.
Теории фильтрации сенсорной информации. Фильтрация сенсорной информации (sensory gating) подразумевает активный процесс (происходящий, например, при участии внимания), посредством которого сенсорные системы отфильтровывают ненужные входящие сенсорные сигналы для облегчения восприятия важных. Соответственно, нарушение этого процесса приводит к наводнению сенсорных систем огромным количеством ненужной информации.
Теории ошибочного приписывания значимости (салиентности). Существует предположение, что аномалии дофаминовой передачи приводят к изменению процессов придания стимулам мотивационной значимости. Объективно нейтральные стимулы (в том числе внутренние: воспоминания или озвученные в голове мысли) приобретают аномальную значимость, что, в свою очередь, требует аномальной интерпретации этих стимулов. Хотя данные теории пока недостаточно четко объясняют собственно феномены нарушения восприятия, элементы этой концепции ошибочного приписывания значимости включались некоторыми исследованиями в модели «сверху-вниз».
Предиктивное кодирование, Байесовские выводы и другие модели «сверху-вниз». В данных моделях нарушения восприятия представляются чрезмерными выражениями нормального процесса перцептивного смещения в зависимости от смещения ожиданий. Особое место занимает модель упреждающего сигнала (corollary-discharge model), согласно которой слуховые галлюцинации представляют собой внутренний голос, ошибочно приписываемый внешнему объекту. Возникает это из-за того, что нарушаются процессы создания эфферентной копии. Этим термином обозначают возникающую в мозге модель предстоящего действия, передающуюся в другие отделы мозга; в норме этот процесс обеспечивает адекватную реакцию на производимое действие (в частности, именно благодаря созданию эфферентной копии невозможно пощекотать самого себя, т. к. чувствительные зоны заранее получают сигнал о том, что это действие совершается самим человеком, и «отключают» защитный рефлекс). Упреждающий импульс также можно считать нисходящим («сверху-вниз») предиктивным сигналом. В других моделях иерархический вывод реализуется при помощи различных алгоритмов развития внутренних ожиданий. В частности, согласно модели циркулярного выведения (circular-inference model), при шизофрении возникают изменения активности интернейронов, приводящие к своеобразной реверберации информации.
Доп. 2. Нейробиологический субстрат процессов принятия зрительных перцептивных решений
Главная и хорошо изученная нейронная сеть, отвечающая за принятие зрительных перцептивных решений включает в себя такие отделы мозга, как средняя височная доля (middle temporal (MT) area, также известная как зрительное поле V5) и латеральная межтеменная кора (lateral intraparietal cortex, LIP) (cм. Рис. 3). Функционирование этой сети было хорошо изучено на обезьянах, выполняющих задания, связанные с движением визуального стимула, например, задание со случайным движением точек (random-dot-motion task). Транзиторные изменения активности нейронов MT кодируют моментальное доказательство движения точки в каком-то из направлений. LIP, получающий нисходящие влияния от MT, наоборот, обладает постоянной активностью, отражающей процесс накопления доказательств, аналогичный таковому в диффузионно-дрейфовой модели. Стоит отметить, что активность нейронов LIP усиливается пропорционально силе накапливаемых сенсорных доказательств (как правило, регулируемой посредством изменений когерентности движений). Изменения, связанные с расчетом предшествующей вероятности движения в ту или иную сторону, ассоциированы с повышенной базальной (престимульной) активностью нейронов MT, «настроенных» на ту или иную сторону, и нейронов LIP. Этот же вывод подкрепляется фМРТ-исследованиями на людях, в которых тоже было показано изменение базальной активности МТ в тех же условиях. Более того, ассоциативные регионы, лежащие иерархически ниже LIP, обладали сходным паттерном активности. К таким регионам относятся лобные доли и ассоциативные зоны стриатума, исходя из чего можно предположить, что процесс накопления доказательств регулируется широкой сетью, затрагивающие как корковые, так и подкорковые структуры.
Таким образом, интеграция предшествующих ожиданий может осуществляться посредством накопления доказательств, обусловленных изменениями базальной активности нейронов, «настроенных» на определенные особенности стимула, которые с большой вероятностью проявятся в данных обстоятельствах. Хотя конкретные зоны, регулирующие эту базальную активность, пока не установлены, имеются данные о роли префронтальной коры, оказывающей нисходящее усиливающее влияние на нижележащие регионы, кодирующие ожидаемые сенсорные стимулы.
Доп. 3. Другие предполагаемые нейромодуляторные системы
Экстрастриарные дофаминовые системы
Кортикальный дофамин. Известно, что высвобождение дофамина в коре при шизофрении снижено, однако пока доподлинно неизвестно, как этот факт связан с развитием перцептивных нарушений. Тем не менее, дофаминергическая модуляция рекуррентной активности кортикальных сетей может играть роль в процессах слуховой рабочей памяти и перцептивными нарушениями. Стимуляция дофаминовых нейронов, чьи отростки идут к слуховой коре, изменяет «настройку» ее нейронов. Соответственно, подобный процесс может внести свой вклад в развитие нарушения восприятия посредством чрезмерного представления характерных для речи особенностей стимула.
Дофамин таламуса. Имеются данные о возможной роли аномальных таламо-кортикальных влияний в развитии нарушений обработки информации при психотических состояниях. Предполагается, что наиболее важны таламо-кортикальные связи с первичной слуховой корой, усиливающиеся при увеличении плотности D2-рецепторов в таламусе.
Холинергическая система
Холинергические системы вовлечены во многие аспекты обработки сенсорной информации. Известно, что у грызунов быстрые колебания уровня ацетилхолина в префронтальной коре связаны с увеличением процента как верного распознавания сигнала в стимуле, так и «ложной тревоги». Стимуляция холинергических базальных отделов переднего мозга у грызунов приводит к реорганизации пластичности в слуховой коре. Математическое моделирование также предполагает роль кортикального ацетилхолина в сигналинге, связанном с фактором (ожидаемой) предшествующей неопределенности при принятии перцептивного решения. Этот вывод подтверждается результатами некоторых фМРТ-исследований на людях.
Тем не менее, конкретная роль ацетилхолина в психотических процессах все еще недостаточна ясна. Некоторыми исследователями предполагается определенная роль ацетилхолина в развитии психоза как при шизофрении, так и при деменции. Клиническая эффективность холинергических препаратов в проведенных на данный момент исследованиях в лучшем случае сомнительна. Холинергическая система и ее связь с развитием психоза все еще требует тщательного изучения, в особенности востребованы исследования, основанные на современных молекулярно-визуализационных методах с использованием недавно разработанных холинергических радиометок. Особенно важными могут оказаться холинергические интернейроны в стриатуме, если принять во внимание их роль в регулировании локального высвобождения дофамина в этой структуре.
Норадреналиновая система
Норадреналиновая система голубого пятна принимает участие в процессах пробуждения и удерживания внимания. Кроме того, предполагается ее участие в модулировании перцептивного решения путем опосредования сигналов о неожиданной неопределенности. К сожалению, на сегодняшний день имеются лишь косвенные и отрывочные сведения о вовлеченности этой системы в развитие психоза.
Автор перевода: Кибитов А.А.
Редакция: Филиппов Д.С.
Источник: Horga G, Abi-Dargham A. An integrative framework for perceptual disturbances in psychosis. Nat Rev Neurosci. 2019 Dec;20(12):763-778. doi: 10.1038/s41583-019-0234-1.