
Функция речи, которая приходит на ум первой – общение, и с точки зрения биологии речь – это коммуникативная система, характерная для представителей рода Человек. С одной стороны, способность к коммуникации распространена в природе повсеместно: в соловьиных песнях, пчелиных танцах, окраске хамелеона и запахе цветов заложена информация, которую важно передать и получить в целях размножения и поддержания жизни. Характерно это и для человеческой речи, но она пронизывает всю деятельность человека и далеко выходит за рамки простого сообщения – то есть филогенетически древний принцип получил у человека такое развитие, что приобрел совершенно новые черты.
Чем человеческая речь отличается от коммуникации у животных?
Наше мышление невозможно представить без языка, и исследования по нейровизулизации показывают, что внутренняя речь (сопутствующая мышлению) задействует во многом те же структуры, что и «внешняя» [1]. У животных дело обстоит иначе: соловей не думает на языке своих песен, а для другого соловья они немногим отличаются от остальных «сигналов извне», например, восхода солнца. Создается впечатление, что животные «понимают» друг друга, поскольку они могут предсказуемо реагировать на определенный сигнал, но говорить так – значит наделять животных человеческими чертами. На самом деле речь идет лишь о нормальном для данной особи ответе на такой сигнал, который не требует обучения и предусмотрен эволюцией.
Интересно, что такой способ общения сохраняется и у человека наравне с языком – это так называемая невербальная коммуникация. Как утверждает общепринятая теория базовых эмоций, через мимику, жестикуляцию и интонации голоса передается наше истинное эмоциональное состояние. Однако в последнее время появилась точка зрения, которая представляет нашу мимику как эгоистичный и манипуляторский прием, в духе многочисленных примеров из животного мира, когда коммуникацией пользуются чтобы обмануть и приманить жертву или партнера [2].
Разницу между речью и невербальной коммуникацией можно держать в голове в качестве примера, сравнивая язык человека и способы коммуникации у животных:
- Коммуникация, обеспечивающая конкретные процессы жизнедеятельности – единственная цель сигналов у животных, но далеко не единственная функция речи у человека.
- У других живых организмов обучение, как минимум, играет меньшую роль в освоении метода коммуникации, он может быть и полностью врожденным.
- Если обучение все же происходит, у животных оно следует иному принципу.
Во-первых, есть какой-то набор сигналов, которые обязательны к освоению, поскольку без них особь просто не выживет. Избыточность здесь недопустима, так как в гонке естественного отбора выиграет вложившийся в охоту или поиски жилья. Человек в этом отношении пользуется свободой: он никогда не осваивает всех слов родного языка, но, в то же время, знает гораздо больше, чем необходимо для удовлетворения элементарных потребностей, поскольку естественный отбор уже не так к нему строг.
Во-вторых, основной способ научения у животных – имитация, им необходимо научиться только одному: точному воспроизведению. У человека, особенно на ранних этапах, имитация тоже важна, но в дальнейшем становится ясно, что важнее усвоить принцип, а потом пользоваться готовыми схемами, чтобы получить множество комбинаций. Как говорит лингвист Светлана Бурлак, ребенок не вызубривает язык наизусть, а достраивает.
- Другие представители животного мира также могут использовать изощренные системы сигналов, в том числе разнящиеся от популяции к популяции, как языки людей. Но для человеческих языков характерна особенная сложность и внутренняя структура. Лингвист Чарльз Хоккет приводит список таких отличительных свойств, которые в совокупности еще и дают определение языка [3]:
– семантичность, то есть соответствие между определенным предметом, явлением окружающего мира и элементом языка, называющим его словом;
– двойное членение, то есть внутренняя структура, в которой значащая речевая единица, с одной стороны, может становиться «кирпичиком» для построения более крупных значащих единиц, а с другой – элементарная смысловая единица может быть разделена только на элементы, уже не имеющие самостоятельного значения.
– иерархичность – сходный принцип, который означает, что речевые единицы могут быть классифицированы от элементарной к более сложным. В системе знаков – начиная от морфемы и заканчивая текстом, в системе устной речи – от фонемы к фонетическому предложению.
– дискретность – четкое различие в звучании (или написании) выражений, имеющих разный смысл, отсутствие плавных, незаметных переходов между ними;
– открытость, то есть возможность создать неограниченное количество сообщений из ограниченного количества структурных единиц;
– культурная преемственность – невозможность впервые усвоить какой-либо язык иначе, чем общаясь с его носителями;
– перемещаемость – принципиальная возможность говорить не только о «здесь и сейчас», а мысленно «перемещаться» во времени и пространстве;
– уклончивость – возможность строить бессмысленные и ложные выражения;
– рефлексивность – возможность с использованием языка рассуждать о нем самом.
Отличия разительные, и некоторые ученые сомневаются, можно ли вообще человеческий язык сравнивать с другими коммуникативными системами и «выводить» из них. Например, лингвист Дерек Бикертон утверждает, что коммуникативная функция для языка вторична, а главная его задача – опосредовать мышление [4].
Как изучают происхождение речи?
В поисках момента возникновения человеческой речи гуманитарные и естественные дисциплины идут навстречу друг другу с разных концов и никак не могут встретиться.
Лингвисты изучают язык ребенка, ситуации, когда зарождается новый язык (например, пиджины и креольские языки), а также сравнивают языки мира, ищут элементарные общие принципы. Однако во всех этих случаях они имеют дело с уже сформировавшейся системой, заложенным принципом, а увидеть его появление возможности нет.
Биологи сталкиваются с другой проблемой: какую бы модель они ни избрали, до языка оказывается еще очень далеко, и отрывочные сведения, собранные на разных животных, не дают представления о зарождении языка. Тем не менее, интересные данные при работе с животными можно получить несколькими способами:
– изучая виды с развитыми системами коммуникации, которые могут в чем-то быть похожи на человеческую; сюда относятся певчие птицы, китообразные или общественные насекомые.
– изучая приматов, ближайших родственников человека: как устроен их мозг, такой близкий по строению к нашему, но не обеспечивающий владение речью? чему возможно научить человекообразных обезьян и какие структуры, гомологичные человеческим, обеспечивают эти способности? какой путь прошла эволюция гоминид, и как менялись необходимые для речи структуры на этом пути?
Есть и еще одна проблема, мешающая продуктивной совместной работе лингвистов и биологов: специалисты в одной области, как правило, недооценивают сложность другой. Лингвисты не всегда понимают, какую строгую проверку естественным отбором должно пройти каждое анатомическое и функциональное образование, необходимое для членораздельной речи. В свою очередь, биологи игнорируют так называемый «парадокс преемственности»: очевидный разрыв между коммуникативными способностями приматов и человека, в то время как прочие изменения происходили плавно и вполне объяснимо.
В какой момент появилась речь, и что было для этого необходимо?
В производство речи вовлечены несколько систем.
Во-первых, нервная; она не только координирует все необходимые движения, но и каким-то образом строит мышление и память на заданной речью основе. Сюда относятся и анализаторы, с помощью которых речь воспринимается.
Во-вторых, часть пищеварительной системы, а именно органы ротовой полости, которые участвуют в артикуляции.
В-третьих, дыхательная система: все мышцы, обеспечивающие вдох и выдох, дыхательные пути, и особенно гортань, необходимая для фонации.
Проследить, как формировались приспособительные адаптации, можно двумя способами. Во-первых, сравнивая, как устроены и функционируют перечисленные системы у человека и у человекообразных обезьян. Как уже говорилось, недостатком этого метода является существенная разница в развитии и невозможность судить о промежуточных этапах. Во-вторых, некоторые данные можно извлечь из сохранившихся останков гоминид. Однако практически все органы перечисленных систем состоят из мягких тканей и не сохраняются, поэтому об их строении можно судить лишь косвенно, по различным следам и отпечаткам, размерам костей.
Познакомимся с данными палеоантропологии, чтобы понять, в какой момент появились известные на сегодня признаки адаптации к речевоспроизведению [6,7].
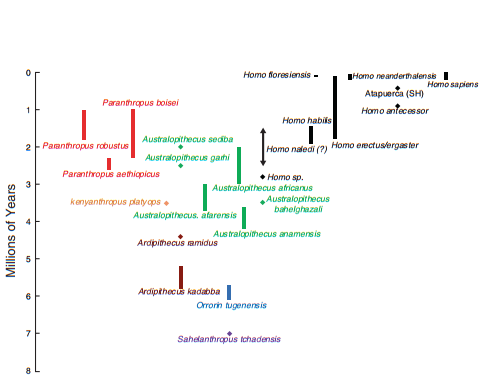
Человеческое эволюционное дерево (Quam, 2017)
- Низкое положение гортани.
Эта анатомическая особенность создает возможность для свободных движений языка, как в горизонтальной, так и в вертикальной полости, то есть для более четко артикулированной речи. В то же время оно обеспечивает возможность издавать более низкие звуки, а поскольку обычно их издают особи более крупного размера, таким образом можно ввести соперников в заблуждение, показаться больше, чем на самом деле. Однако низкое расположение гортани создает и серьезную опасность подавиться: у большинства животных она расположена выше именно для того, чтобы дыхательная и пищеварительная система не пересекались. То есть адаптация, создавшаяся при опускании гортани, должна перевешивать этот риск.
Мнения исследователей, считать ли низкое положение гортани признаком владения речью, разделились. Одни говорят, что одного лишь его недостаточно, чтобы речь создала преимущество. Способность издавать отрывистые крики не равноценна опасности подавиться, значит, основная эволюционная цель опускания гортани у гоминид – создать впечатление о большем размере издающего звук. Эту позицию, например, аргументирует Уильям Фитч [5]. Обратное утверждает, например, Светлана Бурлак [3]: для социальных животных, которые знают и запоминают друг друга, такие способы обмана бесполезны, то есть для опускания гортани должны быть другие причины.
Положение гортани по скелету можно определить лишь приблизительно. Антропологи ориентируются на значение базикраниального угла, однако на современных человекообразных обезъянах и человеке показано, что корреляция между ним и высотой гортани недостаточна. Тем не менее, известно, что у австралопитеков и Homo habilis изгиб основания черепа примерно такой же, как у ныне живущих приматов, а впервые приближенным к уровню современного человека становится у Homo ergaster.
- Подъязычная кость
Подъязычную кость древних гоминид, в отличие от гортани, ученые могут увидеть своими глазами, но часто она находится далеко от места захоронения или вовсе теряется. На редких обнаруженных образцах можно найти разной степени выраженности отметины, свидетельствующие о прикреплении мышц языка и гортани, а также вдавления, к которым прилегают характерные для приматов воздушные мешки.
Судя по подъязычной кости, у Australopithecus afarensis воздушные мешки еще имелись, это мешало бы воспроизведению точных, дискретных звуков. Неандерталец и Homo antecessor (Atapuerca) по строению подъязычной кости ближе к современному человеку.
- Строение нижней челюсти
Подбородочный выступ не является адаптацией сам по себе, но служит ее признаком. Известно, что в эволюции гоминид лицевой череп облегчается, в том числе подвергается редукции верхняя челюсть. Нижней это касается в меньшей степени, в частности, потому что подбородочный выступ служит местом прикрепления многих мышц, участвующих в артикуляции. Однако, некоторое облегчение нижней челюсти способствует ее большей подвижности. Выраженный подбородочный выступ имеется только у неоантропов, у неандертальцев отмечается лишь тенденция к его формированию.
Также интересно исследование рельефа внутренней поверхности нижней челюсти, где у человека имеется подбородочная ость для прикрепления сухожилия подбородочно-язычной мышцы, а у человекообразных обезьян, наоборот, ямка, поскольку та же мышца крепится мясистой частью.
- Контроль дыхания и грудной отдел спинного мозга.
Также для речевоспроизведения необходим произвольный контроль дыхания, в частности способность управлять межреберными мышцами. Эти мышцы управляются спинномозговыми нейронами, расположенными в грудном отделе спинного мозга. Можно предположить, что позвоночный канал, который вмещает этот отдел, будет расширен у особей, которые лучше контролировали дыхательную мускулатуру. Действительно, было обнаружено, что у неандертальца, гейдельбергского человека этот канал так же широк, как у современного человека, но еще у Homo antecessor более узкий.
- Изменение органа слуха
Так же редко, как и подъязычная кость, но от органа слуха сохраняются слуховые косточки. Кроме того, можно оценить размеры слухового канала, проходящего в черепе. Аудиометрическую кривую для ископаемых гоминид строят с помощью компьютерного моделирования.
Мы знаем, что между шимпанзе и человеком существует разница в чувствительности к звуковым частотам. У шимпанзе к частоте 1 кГц чувствительность макисмальная, но к 3 Гц начинает снижаться, и к 5 кГЦ достигает минимума. Человек более чувствителен к частотам от 1 до 5 кГц, которые и соответствуют человеческой речи.
Модификация от состояния слуха шимпанзе к человеческому происходила, по всей видимости, в два этапа. У ранних гоминид (A. africanus, P. robustus) пик чувствительности приходится уже на 1,5-3 кГц. У приматов рода сравнению с предшественниками слегка снижается чувствительность конкретно в этом диапазоне, но общий диапазон повышенной чувствительности расширяется до 1-5 кГц.
- Изменение черепа
Некоторые характеристики черепа могут косвенно свидетельствовать об изменениях мозга, которые необходимы для овладения речью.
– подъязычный нерв и диаметр его канала
Для дифференцированных движений языка необходима более обширная его иннервация, поэтому существует гипотеза, что с развитием речи становится толще подъязычный нерв и шире его канал, который можно непосредственно измерить на ископаемом черепе.
Одни исследователи утверждают, что у представителей рода Homo этот канал имеет абсолютно и относительно больший диаметр, чем у австралопитеков и человекообразных обезьян. Противники этой гипотезы показывают, что толщина канала у человека и шимпанзе варьирует в примерно одинаковых пределах.
– размер мозга и объем черепа
Исследуя череп, можно не только оценить его внутренний объем, но и судить о рельефности мозга по оставшимся вдавлениям. Объем мозга у здорового современного человека составляет от 700 до 2000 см³, и прямой корреляции между когнитивными способностями (также как способностью к речи) и размерами мозга не существует. Тем не менее, это параметр, который сравнительно легко отследить, и в ряду гоминид он заметно изменялся.
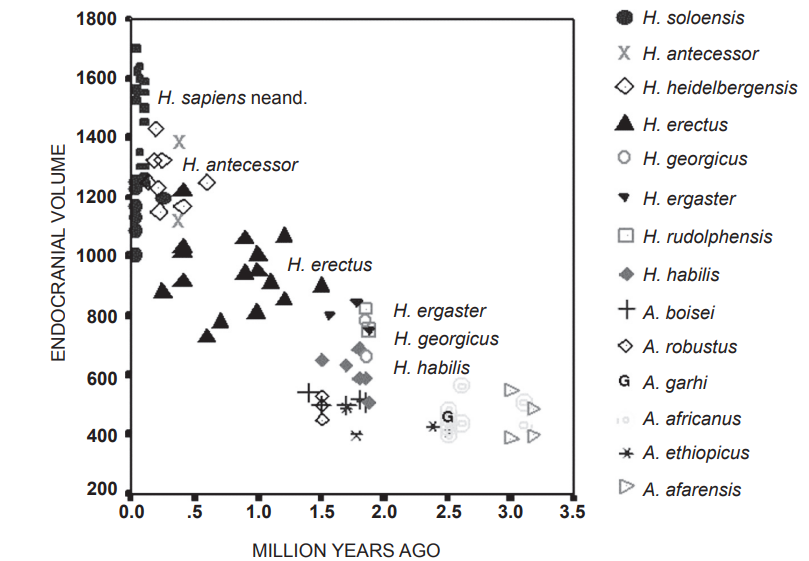
Изменение эндокраниального объёма с течением времени (Holloway, 2015)
У первых представителей рода Homo объем мозга резко возрос и достиг значений, нормальных для современного человека. Однако затем проявилась обратная тенденция, объем мозга снижался, что сопровождалось его структурной перестройкой, в том числе с увеличением рельефности. Видам перестройки, которые не затрагивают объем мозга, на данный момент придается даже большее значение в эволюции человека.
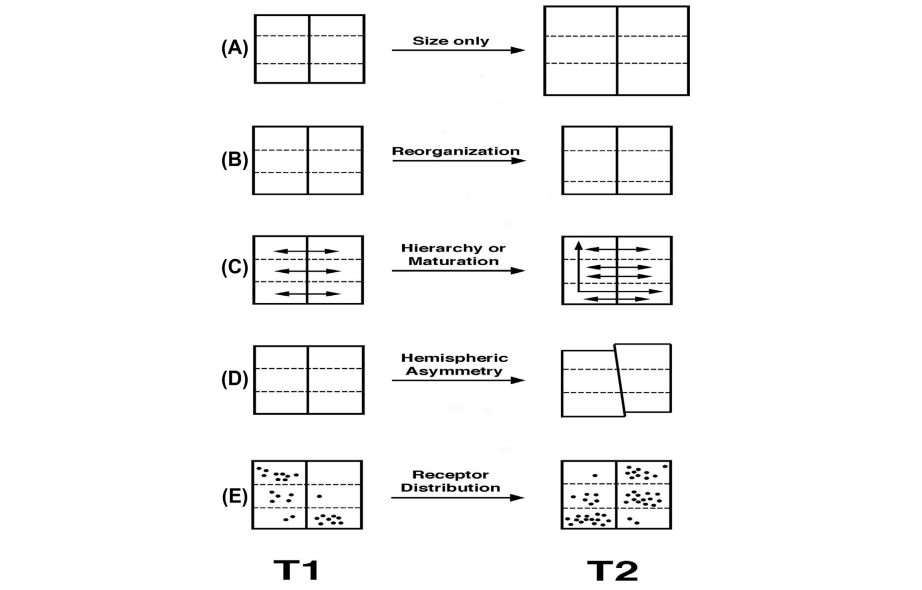
Виды реорганизации без увеличения размера мозга (Holloway, 2015)
– развитие асимметрии [8]
У современного человека участок мозга, традиционно называемый зоной Брока, асимметрично увеличен, чаще в левом полушарии. У человекообразных обезьян такая особенность тоже встречается, но является редкостью. Способ определения увеличенной зоны Брока по вдавлениям на черепе также вызывает вопросы, однако такие вдавления обнаруживаются у представителей рода Homo (H. rudolfensis, H. erectus, H. heidelbergensis, H. neanderthalensis) и не обнаруживаются у австралопитека.
Еще один признак асимметрии – так называемые petalias, неравномерности полушарий, когда одна из долей больше по объему и, возможно, переходит через срединную линию, несколько оттесняя аналогичную ей с другой стороны. Такие особенности у человека изучены, например, у правши левая затылочная и правая лобная доли, как правило, несколько больше коллатеральных. Длительное время считалось, что они есть только у человека и в эволюции появились у представителей рода Homo, однако в последнее время было показано, что они встречаются и у других высших приматов.
Что все это время происходило с мозгом?
Ответа на этот вопрос палеонтологические находки дать не могут. На данный момент «живой» мозг доступен исследованию только в двух точках: условном начале пути, то есть на примере наших ближайших родственников-приматов, и в конце пути – собственно, у человека.
История начинается с вопроса: почему именно приматы? Это не единственные социальные организмы на Земле, высокие когнитивные способности (одновременно с развитыми коммуникативными системами) отмечаются у китообразных, некоторых видов птиц. Часто говорят, что все в эволюции – непредсказуемое совпадение большого количества факторов. Попробуем перечислить факторы, которые могли способствовать появлению речи именно у приматов [5, 9]:
– условия жизни
Приматы – социальные животные, которым коммуникация необходима в том или ином виде, и с усложнением деятельности она приносила бы все большую пользу. Кроме того, значение имеет размер группы, в которой состоит особь: было показано, что проживание большими группами – само по себе вызов для мозга и способствует его развитию. Как пример приводят способность присваивать другим особям примитивные «имена» и обмениваться некой информацией о них — она облегчила бы облегчила бы кооперацию в группе.
– рабочая память
Традиционно считается, что у животных возможности рабочей памяти очень ограничены: они не удерживают сенсорную информацию больше нескольких секунд, легко поддаются интерференции и не обнаруживают механизмов повторения (rehearsal) зрительно-пространственной или фонологической информации, которые у человека постоянно активны. Все это несовместимо с полноценной речью, поскольку речь состоит еще и в манипуляции объектами, содержащимися в памяти. Тем не менее, у приматов (а также у врановых) обнаруживают отдельные действия, которые хочется назвать запланированными: например, одна содержащаяся в неволе шимпанзе, несмотря на препятствия со стороны сотрудников зоопарка, каждый раз заранее набирала кучу камней, чтобы бросаться ими в посетителей зоопарка.
Структуры, ответственные за поддержание рабочей памяти, были первоначально обнаружены у людей, переживших инсульт. Однако затем эксперименты начали проводить на приматах, как имеющих максимально схожую с человеческой нервную систему. К тому же с простыми заданиями, требующими выбрать недавно показанную картинку, обезьяны справлялись неплохо. Именно на животных есть возможность применять наиболее точные методы, связанные с регистрацией активности единичных нейронов. Этим способом было показано, что существует небольшая область, которая активна при пассивном восприятии стимула (любой модальности) и при его удержании «до востребования» – то есть, вероятно, важная именно для рабочей памяти. Эта область кодируется как Spt, от Sylvian, parietal, temporal. И она хорошо известна афазиологам: в исследованиях, которые пытались конкретизировать область повреждения, соответствующую симптомокомплексу афазии Брока, у всех пациентов участки повреждения пересекались именно в ней.
– гомологичные структуры
Выяснив, что ключевую функцию в процессах рабочей памяти играет префронтальная кора, ученые занялись ее исследованием. Если раньше считалось, что единственное место, где может «храниться» память – это сенсорная кора, теперь появилась возможность исследовать память, как принцип. Первоначально работа велась на примере зрительной системы: было показано, что существуют два разных пути, связывающих зрительную кору с разными участками префронтальной: один из них связан с обработкой информации о пространственных характеристиках, другой – о характеристиках лиц и объектов (дорсальный и вентральный потоки). Именно к этой информации обратились исследователи речи, когда были обнаружены гомологи зон Брока и Вернике у приматов, и необходимо было определить, как формировалась связь между ними.
Участки, гомологичные зонам Брока и Вернике, у приматов редуцированы. Что привело к их развитию? Две области, гомологичные зоне Брока у приматов, получают информацию, связанную с восприятием звука. Область 44, «сенсорная», – по вентральному потоку, а точнее от вентролатеральной префронтальной области, где сходятся вентральные потоки зрительной и слуховой информации. Область 45, «моторная», получает информацию по дорсальному потоку, в том числе и от области Spt, связанной с нижними отделами теменной доли, которая у человека задействована в звуковом аспекте рабочей памяти.
Основываясь на этих сведениях, выдвинули гипотезу о первичной роли языка: в какой-то момент приматам пригодилась способность называть предмет, не присутствующий сейчас в поле зрения, что требует определенных характеристик рабочей памяти. Развитие зоны Брока происходило за счет развития связей в нескольких направлениях одновременно. Во-первых, с передними отделами теменной доли (через вентральный поток), где обрабатывается информация о внешнем виде предмета (или особи), который нужно назвать. Во-вторых, с нижними отделами теменной доли, что позволяло удерживать в памяти звуки и соотносить с действиями артикуляционного аппарата. Однако аркуатный пучок, наиболее известный проводящий путь между зонами Брока и Вернике, у приматов не выражен. Ему соответствует небольшой пучок волокон, соединяющих область 45 и зону Tpt, предположительный гомолог зоны Вернике. Также слабо развиты у приматов собственные связи зоны Вернике, например, с нижними отделами теменной коры.
– контроль над вокализацией
Нейронные механизмы контроля организованы в трехуровневую систему. Первый уровень – так называемое шасси ствола мозга. Оно включает комплекс ядер ствола, контролирующих движения челюстей, губ и гортани (тройничный, подъязычный, блуждающий), а также внутренние структуры ствола (например, ретикулярная формация). Второй уровень – подкорковые центры, среди них особую роль играет околоводопроводное серое вещество. По всей видимости, для простой, рефлекторной вокализации управление более высокого порядка не является необходимым. Третий уровень – корковые системы. Одна из них более примитивна и имеется у всех млекопитающих – это срединная кортикальная система, которая включает поясную кору и прилежащие области. Она участвует в волевом контроле воспроизведения и прекращения вокализации, в том числе и у человека, и ее действие касается не только речи, но и таких непроизвольных форм, как, например, смех. Однако в эволюционном плане нас более интересует латеральная кортикальная система. У человека она непосредственно связана с моторными нейронами ствола (хотя опосредованная тоже существует), и разрушение корковых центров приводит к афазии. У других млекопитающих, в том числе обезьян, связь между корой и ядрами ствола непрямая, и даже анэнцефалия не препятствует врожденной вокализации.
– зеркальные нейроны
Так называемые зеркальные нейроны, которые участвуют в «переводе» наблюдаемых действий на язык внутренних моторных программ, обнаруживаются во многих областях, связанных с речью. Например, на границе с зоной Вернике, в области дорсальной премоторной или первичной моторной коры. Однако отчетливое участие этих нейронов в речевой активности обнаружено только у человека, у обезьян оно ограничивается причмокиванием и другими движениями лицевой мускулатуры. Возможно, в эволюции их значение возрастало. Нейроны с такими функциями могли бы играть значительную роль в имитации жестов, вокальных сигналов, таким образом облегчая научение.
– функциональная латерализация
У обезьян обнаруживаются признаки как функциональной, так и анатомической латерализации, но в меньшей степени, чем у человека. При этом структуры, которых касается латерализация, часто относятся именно к коммуникации (височная площадка, связи гомолога зоны Брока). Однако в эволюции должны были присутствовать дополнительные факторы, способствующие углублению латерализации. Одно из очевидных проявлений латерализации – тенденция пользоваться правой рукой для определенных действий, часто требующих тренировки. У обезьян она не так развита, как у человека, но проявляется, когда требуется соорудить простое приспособление или что-то бросить. Возможно, способность к созданию орудий, которая развивалась в ряду гоминид и требовала латерализации, подтолкнула к доминированию левое полушарие, которое изначально несколько специализировано для целей коммуникации, и способствовала большему его развитию.
Подготовил: Шишковская Т.И.
Источники:
- Geva S. et al. The neural correlates of inner speech defined by voxel-based lesion–symptom mapping //Brain. – 2011. – Т. 134. – №. 10. – С. 3071-3082.
- Crivelli C., Fridlund A. J. Facial displays are tools for social influence //Trends in cognitive sciences. – 2018.
- Светлана Бурлак, Происхождение языка: Факты, исследования, гипотезы
- Bickerton D. Species and language. – 1990.
- Фитч У. Т. Эволюция языка. М.: Языки славянской культуры, 2013. 768 С.
- Бурлак С. А. Время появления звучащей речи по данным антропологии // Вестник Московского университета. Серия 23: Антропология. — 2012.
- Quam R. M. et al. Evolution of Hearing and Language in Fossil Hominins //Primate Hearing and Communication. – Springer, Cham, 2017. – С. 201-231.
- The Oxford Handbook of Language Evolution, Tallerman and Gibson
- Francisco Aboitiz – A Brain for Speech: A view from Evolutionary Neuroanatomy – Palgrave Macmillan UK 2017